

Journal of Creation 10(2):219–240, August 1996
Browse our latest digital issue Subscribe
Australopithecus and Homo habilis—pre-human ancestors?
A survey of the palaeoanthropological literature reveals the controversies raging between various discoverers of australopithecine and habiline bones and their followers. It can also be readily demonstrated that preconceptions have already decided the interpretation of these bones as belonging to human ancestors, even when the contrary evidence is obvious. Debate continues over the australopithecines, but close examination of the dentition and jaws, the position of the foramen magnum, the upper body bones, the rib-cage and waist, the arm, hand and phalanges, the pelvis, hip and thigh, the legs, knees and feet, and the ankle joint not only shows that they were not bipedal, but that they were probably the ancestors of today’s great apes, the chimpanzees and gorillas. The habilines likewise have been difficult for the palaeoanthropologists to classify as a uniform group, because the evidence clearly shows that they were variants of the australopithecines and not a separate taxon within the genus Homo. Only evolutionary bias has hindered them from concluding that apes and humans are unrelated and were created separately.
Introduction
Various camps in the field of human historical study have manipulated or distorted (probably unconsciously) the facts about the australopithecines and habilines to suit their own particular viewpoints, and consequently their own fame, fortune and standing in the scientific community, plus the very generous taxpayer-funded research grants available.
Not only are a number of experts at variance with each other, but most have allowed their preconceptions to colour facts to such an extent that an entirely false picture of human origins now dominates the entire world’s educational systems and public media networks. There is no scientific reason for insisting that any of the australopithecines and habilines are ancestral to humans, though they may be the (hitherto missing) non-evolutionary forbears of modern great apes.
[It should be made perfectly clear at the outset, that when evolutionist authorities are cited as being critical of various aspects of the human evolutionary ‘structure’, it is not implied that they now reject the theory. All are still committed evolutionists, despite the heated nature of their disagreements: in fact, they quickly close ranks and temporarily forget their disunity when threatened by creationist criticism.]
Ever since the discovery of the first Neanderthal remains in the 1850s, many palaeoanthropologists have displayed excessive zeal in depicting human origins as being the result of a blind and purposeless process which caused some (as yet still unknown) form of ape to transmute into modern men and women.
The most important stage in this chain of alleged events was the transformation of a quadrupedal chimp-like ‘common ancestor’ into a supposedly erect group of creatures described as australopithecines some 4 to 5 million years ago (Ma). These curious animals, mostly discovered since 1924 in various regions of southern and eastern Africa, have become the only candidates for the alleged transition to ‘primitive’ man. Another protohuman which supposedly links these creatures to humans is the so-called habiline group, which supposedly arose after the gracile australopithecine stage around 2 Ma, and which thereafter evolved directly into Homo erectus approximately 1.8—1.9 Ma.
These putative events, spanning a period of about 3 million years from mid-Pliocene to early Pleistocene, form the foundation of evolutionary theory in respect of human beings.
However, almost every single fossil link in this ‘chain’ can be successfully challenged by educated and informed creationists by demonstrating the biased interpretation of much of the evidence by the experts. In truth, there is often precious little science but much speculation; this contention is powerfully supported by recent publications from within scientific ranks.
| Humans | Apes |
|---|---|
| Large cranial capacity | Small cranial capacity |
| Flattish face | Prognathous face |
| Short arms, long legs | Long arms, short legs |
| Anterior nasal spine | Nasal spine absent |
| Generally steep forehead | Little or no forehead |
| Small canines | Large canines |
| High rounded cranial vault | Low cranial vault |
| No diastemata | Diastemata present |
| Little body hair | Thick body hair |
| Bipedal locomotion | Quadrupedal locomotion (usually) |
Table 1. Some of the main differences between the morphology of apes and of humans. Despite a similar molecular structure, there is a huge gap between man and the apes. It should be noted that some humans such as erectus and neanderthals have a degree of facial prognathicity, but not to the degree of the apes/australopithecines. Cranial capacity of humans generally ranges from about 900–1400 cc, while the ape/australopithecine range is from about 350–750 cc.
Roger Lewin is a highly respected palaeontologist and palaeoanthropologist, but upon observing the antics of his fellows over the past few decades, he felt compelled to expose the bitter divides, personal ego-trips, jealousies, and hunger for fame of many workers in the field who have dominated these branches of science in this century.1 Lewin himself still remains committed to an evolutionary origin for humanity, despite his clear view that there has been far too much emotionalism and preconception on the part of researchers in their quest.
Creationists, while accepting and welcoming the type of criticism and frankness displayed by Lewin, can go further by drawing attention to the underlying weaknesses and contradictions in the naturalistic position, the often vacuous arguments used in palaeoanthropological circles, and the many flaws in the theory.
It is proposed to reinforce Lewin’s criticisms by examining the contested evidence surrounding human fossil remains; that is, those bones and teeth whose significance is disputed by various individuals and factions within the scientific community. Areas which are not contested (such as the extremely ape-like crania of the ‘protohumans’) will not be generally discussed in any detail, because there is little point in arguing over items which are accepted by all, including creationists.
Any human phylogeny (evolutionary family tree) requires fossils to fill the large gap between extant apes and man (see Table 1). It would be expected that such fossils should represent bipedal ape-like creatures with morphologies intermediate as a whole and in the various individual traits. Such transitional morphology appears, but only at first glance, to occur in a group of ape-like creatures which is outlined below.
It should be pointed out that before the discovery of the australopithecines, a suitable intermediate was expected to be large-brained and modern-looking, but after the australopithecines became gradually and grudgingly accepted in the 1960s, the prediction was clearly on the wrong track—what was found was a small-brained, very ape-like creature. In the early 20th century, conditioned by ‘Piltdown Man’ (which at that time was not known to be a hoax), the missing link was expected to be a quite advanced and more human-like individual. This attitude played an important role in the initial rejection of the Taung Child (a gracile australopithecine), because it was seen at that time to be in conflict with expectations.
The australopithecine subfamily
The first discovery, the so-called Taung Child in 1924 by Raymond Dart, drew little initial interest and was rejected by most experts, but later finds over the ensuing years have revealed a number of similar-type fossils.
The present classification of the subfamily is as follows:
- Australopithecus africanus (the gracile forms including Taung),
- A. robustus (a more heavily built and coarser form),
- A. boisei (a much coarser form of robustus), and
- A. afarensis (found in 1973–1974 in Hadar, Africa), which is believed to be the ancestor of all later australopithecine forms and of humans. Some authorities believe it is not a valid taxon, and should be included as only a sub-species of A. africanus.2,3
(In the last four to five years new fossil finds have been made in Ethiopia, and these will be briefly discussed later).
Just about all of the above types have been proposed as the original human ancestor, but in recent times most experts have plumped for the last afarensis, otherwise known as Lucy, or Lucy/afarensis from Hadar, in Ethiopia (Africa).
We will look at various australopithecine features about which the experts disagree, either mildly or vehemently. Some features are irrelevant to our discussion because all authorities, creationist or evolutionist, agree as to their significance. There is little dispute over such characters as the skull morphology, the long, ape-type arms and the short legs. These fossil parts are clearly pongid and can be put to one side. The features under examination are:
- The dentition and jaws
- The position of the foramen magnum (the opening joining the spinal column to the skull).
- The upper body
- The rib-cage and waist
- The arm and hand, the phalanges
- The pelvis, hip and thigh
- The legs, knee and feet
- The talus (ankle joint)
The question of locomotion—bipedality versus quadrupedality—is also examined in the context of the relevant bones.
In every one of the above features we find varying degrees of dispute, some of which involve the reconstruction of the skull from the fossil remains. Such reconstruction is often influenced by the preconceptions of the discoverer, because he/she can all too easily read their own bias into how the assembled skull or skeleton ‘should’ look.
There are two main or basic opposing camps in paleoanthropology—the Richard Leakey group and the Donald Johanson faction. These two, one British and the other American, have made big headlines in the last 30-odd years because many of the most famous finds have come to light through their field work in Africa. Richard Leakey was the discoverer of the ER 1470 skull in 1972, and of the so-called Turkana ‘Boy’ (a Homo erectus form) in 1984. Johanson made the headlines in the 1970s with his sensational find of the A. afarensis specimen known as Lucy, along with the ‘First Family’ at Hadar. Further afarensis type fossil discoveries have been made in the last couple of years.
Each of the above ‘stars’ has gathered a following of supporters over the years, along with opponents and skeptics. Ranged alongside Leakey are such figures as his mother Mary, and the anatomist Alan Walker. Johanson has been influenced by Tim White of Berkeley, a close colleague. It is not intended to give the impression that the supporting groups always back up the main players, but generally it is true to say that they find themselves in agreement with the ‘stars’. In addition, there are a considerable number of independent researchers, such as Coppens, Lovejoy, Tobias, Wolpoff, Susman, Stern, Olson, Prost, Boaz, Schmid, Tuttle and Spoor, who often find themselves in disagreement with the published conclusions of Leakey or Johanson.
Opposing viewpoints
The Leakey group generally has supported the contention that the genus Homo is very ancient, and is not closely connected to the australopithecines, which they generally regard as being more like close cousins of man. So far, the ‘real’ Homo ancestor is still missing. Richard Leakey’s father, Louis, maintained until his death in the early 1970s, that the Homo line would go back a very long way,4 and naturally his son Richard was overjoyed in 1972 with his discovery of the ER 1470 skull, at that time seeming to reinforce his views. It was only natural therefore that when Johanson made his find of Lucy and the ‘First Family’ shortly thereafter, the discovery was not exactly welcomed by the Leakey camp. Lucy revealed herself to be extremely ‘primitive’ (ape-like), which did not fit into the Leakey pattern.
Since then there has been a persistent feud between the two camps, which has sometimes degenerated into public brawling. The Johanson camp believes that Lucy/afarensis is very close to the alleged branching away from the apes, and they have maintained this general opinion ever since. But what one camp sees as an indication of human ancestry, the other sees as not being significant, although the two groups are now closer in their outlook than they were originally. For example, both Leakey and Johanson maintain that Lucy/afarensis walked fully upright—a view that has been strongly contested by other independent researchers such as Susman and Stern,5 and Spoor.6
As we proceed we shall find many more examples of controversy and disagreement regarding the various fossil remains. Most authorities believe the ‘chain’ of human evolution consists of the lineage A. afarensis to A. africanus to H. habilis to early Homo (H. ergaster or H. erectus), with A. robustus and A. boisei being relegated to a deadend side branch (for example, Clarke, as cited by Armstrong7), but according to Andrews there is a lack of evidence linking the gracile forms and Homo.8 In fact, Andrews goes even further, and says that none of the four australopithecine species are on the line leading to humans.9 In Bromage’s view neither the gracile nor the robust forms are directly on the human line,10 despite the supposed sharing of some characters in early Homo and robustus forms (sometimes known as Paranthropus), but he also believes that skull ER 1470 (a habiline in favour with most authorities) has a “provocative resemblance” to WR 17000, the most hyper-robust, most ape-like specimen ever discovered.11 This will become clearer as we proceed.
Discussion
(1) Dentition and jaws of australopithecines
Lucy/afarensis
It can be clearly seen from the various textbooks that the mandible is V-shaped and ape-like, and lacks the more rounded morphology of humans. Also noted is the rectangular arcade of the palate, as opposed to the parabolic arcade of humans. Apes usually also display the diastema, or gap adjacent to the canines, a feature lacking in humans (but present in afarensis) because our canines or eye teeth are in occlusion—that is, the upper and lower teeth meet fairly evenly and there is no need for a gap to accommodate projecting canines.
Johanson and White argued about the significance of these traits, until White eventually convinced Johanson that the teeth and jaws of afarensis were compatible with other specimens of the first family. Yet Johanson himself had originally been convinced that Lucy’s mandible was the wrong V-shape instead of the parabolic shape of humans. He said plainly: “She was clearly not a human”.12 Gowlett also made reference to this subject:
“The jaws of A. afarensis show a basically human arrangement of the teeth, but the diastema is still very visible in the upper jaw. The U-shape, with the two rows of molars running parallel, also distinguishes the finds from later man.”13
His mention of a basically human arrangement of the teeth is misleading because all apes and humans share this type of arrangement—32 teeth with each jaw containing six molars, four premolars, four incisors, and two canines, plus the Y-5 cusp pattern on the crowns of the cheek teeth. This is not necessarily a sign of common ancestry. Australopithecines also had very large grinding molars.
Tanner said that the overall similarities of jaw and tooth proportions between afarensis and pygmy chimps (bonobos) were “striking”.14
A new afarensis mandible (Maka, VP-1) has been unearthed which displays much the same extremely chimp-like characteristics,15 but there seems to be little basis to reach any firm conclusion because the sample is small, and has not yet been subject to detailed critique by other scientists.
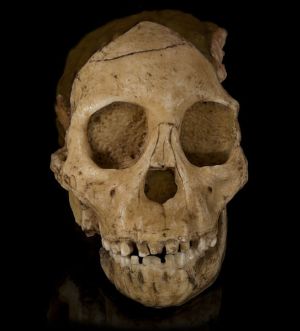
The Taung Child (Type fossil for the category A. africanus) (see Figure 1)

Although most experts claim that this specimen displays an at least partly human dental pattern, there has been much controversy. As an infant of only about three years of age, we are dealing with milk teeth, and not the permanent ones. Because of this, the canines do not project above the occluding surfaces of the other teeth, and Taung therefore lacks the diastema which is present in older apes. Until the canines reach their full length and interlock, there is no need for the gap which accommodates the eye teeth. This lack of a diastema appears to have misled some experts in their haste to find a human feature. Over the years various parties have challenged and re-challenged the significance of the Taung (africanus) dental traits. For instance, Conroy and Vannier commented that:
“… the permanent incisors of the chimp and Taung skull are virtually identical”.16
According to Beynon and Dean, these early ‘hominids’ possessed unique patterns of dental development differing not only from man and apes, but also from one hominid taxon to another.17
It should also be noted that Bromage found the age of the Taung fossil at death was 3.3 years, which is precisely what one would expect for an ape whose first molar is erupting:
“It most definitely was not what you’d expect in a human child with the first molar just emerging [emphasis added].”18
Leakey and Lewin make the further observation that:
“All the australopithecines developed their dentitions quickly, like apes … Bromage found that australopithecine faces are built like ape faces, not human faces [emphasis added].”19
The same authors also cite Conroy and Vannier, who found that the development patterns of Taung are “… very much like those of a 3 to 4 year-old great ape … The evidence is beginning to become overwhelming… early hominids (australopithecines) do appear to have had a more ape-like dental maturation period”, but they then also point to the lack of brow-ridges and the lack of the diastema which indicated to them that Taung was not a little human, but also not a little ape.20 Have they forgotten that all infant apes lack large brow-ridges and diastemata?
A few years ago vigorous debate occurred between those who believed the Taung dentition was little different from an ape’s, and those who thought they could see a more human-like pattern.21–25
Bromage was surprised to find, during his examination of this fossil, that the Taung face and jaws jutted forward like apes, and this will be referred to later.26 Today the clashes seem to have died away, but the volatile nature of the debate indicates that each side saw what they wanted to see—that is, what they expected to see according to their preference or their personal beliefs about Taung.
A good example of emotionalism in the study of human origins can be seen when the Taung discoverer, Raymond Dart, was very surprised to see Scottish-born palaeontologist Robert Broom walk past him at a medical school and drop to his knees in front of the Taung fossil—“Broom said it was ‘in adoration of our ancestors’.”27
(2) The position of the foramen magnum
This is the scientific name for the opening in the underside of the skull where the spinal cord is attached. In apes the opening is more towards the rear of the skull and ‘points’ downward and backward. In humans it faces more downward than backward, thus reflecting the fully upright stance of humans where the head is perched on top of the spine.
In the alleged man-like australopithecines, the position of the foramen magnum varies to some degree. In the Taung child, for instance, which is an infant gracile australopithecine, it appears to face more down than is usual with apes. However, Gish, a creationist, points out that in foetal and infantile stages of apes, the foramen magnum lies relatively further forward but ‘moves’ towards the rear during post-infantile growth, whereas in man the relative position of the structure changes very little during growth. He says: “… it would be a serious mistake in this respect, to compare a juvenile ape skull to an adult human skull.”28
In the infant Taung specimen, the face is flatter and the forehead higher, in similar fashion to all young apes. As the infant grows to adulthood, the face becomes more prognathous, the supraorbital ridges develop, and the forehead becomes flatter. This means that the distance between the foramen magnum and the forward extremity of the jaws increases (unlike humans), while the distance from the skull’s posterior extremity to the foramen magnum remains little changed. The opening itself does not move—all the ‘movement’ is in the lengthening of the skull and jaws as the specimen grows older.
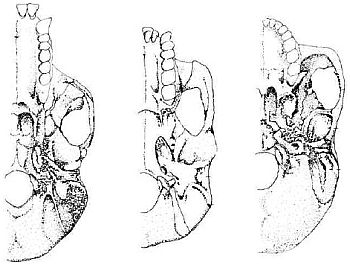
In the illustration of the afarensis specimen reproduced by Johanson and Edey, we see that the foramen magnum lies on the underside of the skull, but still near the chimpanzee position.29 The same illustration shows that the human foramen magnum lies further forward than the chimp or afarensis.30 The distance between the opening and the anterior extremity of the jaws and teeth, in both chimp and afarensis, is much greater than the same measurement in humans (see Figure 2).31 The reason for this is that as the australopithecines (and the chimp) mature, the brain capacity hardly grows at all while the face and jaws become increasingly prognathous.
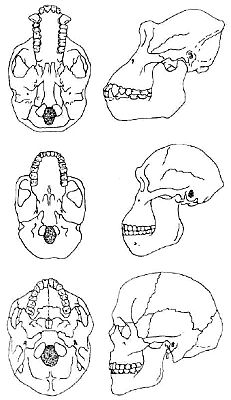
When we compare the skull of the common chimp Pan troglodytes and an adult australopithecine specimen, the foramen magnum lies in much the same position. This is confirmed by Johanson and Edey’s reproduction of the undersides of chimp, human, and australopithecine skulls as shown in Figure 2. In both cases, chimp and australopithecine, the ratio of the distance from the back of the skull to the opening, as compared to the distance from the opening to the anterior extremity of the jaws, is remarkably similar, but in humans the ratio is very different. Therefore, to claim that on this feature alone the afarensis specimen locomoted fully erect in the human manner is very misleading. The human-type foramen magnum position, on the other hand, clearly reflects our erect bipedal locomotion. We see the same situation in the illustration provided by Campbell, which compares the skull undersides of a female gorilla, A. africanus, and a Homo sapiens (see Figure 3).32 There is only a slight difference between the australopithecine and the gorilla, although Campbell claims the africanus position is intermediate. When we allow for the size difference, the gorilla and the australopithecine are almost identical in this respect, and in any case, the measurement ratios plainly show that africanus (supposedly the australopithecine-type closest to humans) lies squarely in the ape range.
In the case of the robustus specimen OH 5 (Zinjanthropus) there is no guarantee that the original reconstruction of the skull was correct, that is, a true reflection of how it looked in real life. The rear portion of this cranium was not physically joined to the facial region, and if the rear half of the cranium is pivoted, we get a very different picture from that of the original Leakey work. Fossil skull reconstruction depends heavily on getting the angle of the jaw/cranium right, but there is simply no way of telling which reconstruction is correct, as these creatures became extinct long ago. If a palaeontologist already believed that australopithecines possessed an upright stance, then this will influence his reconstruction, and the result will be that he would give it a flatter face.
As Willis says:
“We can all see a bone and know it is a bone, but what it is evidence for depends on one’s interpretations [emphasis added]”.33
If one is a convinced evolutionist, one will see fossils in a different light from a non-believer.
In conclusion, the position of the foramen magnum in the australopithecine group is not as favourable to evolution as we have been led to believe, and palaeontologists have read into these fossils far more than is warranted. This conclusion is supported by further evidence from the postcranial remains, as shall be seen later.
(3) The upper body (the shoulder)
A feature not picked up by Johanson, the discoverer of Lucylafarensis, was that in contrast to humans her shoulder joint is cranially-orientated. In modern humans it faces outwards, parallel to the ground, whereas apes have a socket which faces mainly upwards. This fact was revealed by Cherfas, who immediately realised the implications (see Figure 4).34
Attention was also recently drawn to this feature by Richard Leakey and Lewin.35 The authors say that Schmid of the Anthropological Institute of Zurich pointed out that, because the shoulders are important for arm swinging and balance, this chimp-like feature does not seem compatible with efficient bipedality. Cherfas in his report stated that the ape-type afarensis shoulder
“… would certainly be valuable if the arm were held overhead much of the time, as it is when climbing and hanging in trees.”36
I have not seen any reference by Johanson to this problem, which should have been obvious to such an experienced palaeontologist soon after the Afar skeleton was first assembled.
(4) The rib cage and waist
Here is another typical pongid feature, missed by Johanson and his colleagues. He makes no mention of these vital characteristics in his 1990 work Lucy’s Child, co-authored with Shreeve,37 although he vigorously contests other criticism of his ‘Lucy’ skeleton.
Leakey and Lewin again refer to Schmid’s findings:
“The chest was the problem. I noticed that the ribs were more round in cross-section, more like what you see in apes. Human ribs are flatter … but the shape of the rib cage itself (in Lucy) was the biggest surprise of all. The human rib cage is barrel-shaped, and I just couldn’t get Lucy’s ribs to fit this kind of shape. But I could get them to make a conical-shaped rib cage, like what you see in apes”.38
These aspects are also referred to by Zihlman, who points out that the new reconstruction of Lucy ‘… reveals her to be remarkably chimp-like, particularly in the morphology of the rib cage [emphasis added]”.39
But according to Leakey and Lewin, Schmid went even further:
“He examined the whole trunk, the lumbar region, and the shoulders. (All of these) are important in human running; the shoulders for arm-swinging and balance, the trunk for balance and breathing, and the waist for flexibility and swinging of the hips. What you see in Australopithecus is not what you’d want in an efficient bipedal running animal… the shoulders were high, and combined with the funnel-shaped chest, would have made arm-swinging improbable in the human sense. It wouldn’t be able to lift its thorax for the kind of deep breathing that we do … The abdomen was potbellied, and there was no waist [emphasis added].”40
(If you visit a zoo, you will notice the potbellied look of many common chimps, orang-utans, and even gorillas.)
Now Johanson and his colleagues have always maintained that Lucy and all australopithecines were capable of a fully erect, human-type striding gait, and Lovejoy went so far as to say that Lucy was “… even better designed for bipedality than we are”!41
So, how can the various experts be so diametrically opposed in their viewpoints, when they all see exactly the same fossil material? The only answer seems to be that there is often a lack of objectivity by those who have something to gain, whether it be fame or fortune, and therefore it is wise to pay more attention to those experts without any axes to grind, such as Schmid.
(5) The arm and hand
So far as I know, nobody has contested the ape-like dimensions of the humeri of australopithecines, but Cherfas made the passing comment: “Lucy, from Ethiopia, with her ape skull and human body”,42 thus implying that the only doubts about her status were from the neck up. This statement is entirely misleading, because Lucy/afarensis definitely did not possess a human body. Feature after feature turns out to be decidedly non-human, that is, ape-like.
In the case of the long bones of the arm, the simian nature is again clearly evident. Not even the most biased evolutionist would deny that Lucy’s arms dangled down to her knees or lower, but later a so-called Homo habilis fossil (OH 62) was found which clearly proves that a million-year-later specimen still possessed this very pongid feature. One would expect that by 1.4 million years after Lucy’s time, such an ape-type characteristic would long before have evolved into a more human-like arm. Let Leakey and Lewin do the talking43—they say that a key clue lies in the so-called humerofemoral index; that is, the length of the humerus (upper arm bone), when compared with the length of the femur (thigh bone). In humans the index is about 70 per cent (the humerus is 70 per cent of the length of the femur). The figure for chimps is around 100 per cent—that is, both bones are about the same length, which means that in apes, the arms dangle down to the knee and below because of the short legs.
In Lucy’s case the index is about 90 per cent. When we come to her supposedly more advanced descendant, OH 62, an alleged habiline, we find a humerus even longer than Lucy’s—about 264 mm. This means an index of 95 per cent, and as the authors admit, this makes the mistitled ‘H. habilis’ even closer to the chimp! This has proved a nasty problem, as Leakey and Lewin concede.44 Referring to this puzzle, Reader expressed some surprise: “This meant that the primitive ape-like characteristics of A. afarensis must have remained a distinctive and unchanging feature of the ancestral hominid for a very long time. The earliest humans (the so-called habilines) were more like apes than had been supposed [emphasis added]”, and on the same page he showed more surprise that the essential transition from ape to human must have been completed in (only) 200,000 years.45
This unlikely proposition led him to say on the next page: “[This] has left Homo habilis more insecure than it ever was.”
Not only did this discovery put the status of all australopithecines into question, but it also severely damaged the validity of the taxon H. habilis, which now seems to be just a large-brained australopithecine, which in turn means ape.
Johanson and Shreeve could not escape the implications either, although they sought to find a way out by suggesting that two hominids might be involved.46 They gave up this hope shortly after and changed the subject, but returned to the problem later in their book,47 claiming that they could not accurately establish the actual length of the humerus. The problem remains unsolved.
A recent report by Bunney covers new finds of arm and other bones in the Hadar region, which “… complete the image of a waddling, forest-dwelling creature with long, powerful arms, the females lighter and more agile than the males”; and “… early hominids may not have been efficient walkers … Lucy had hips that were wide for their size … [this] would have been less suitable for sustained walking and running.”48
When we turn to hand and phalanges (fingers), once again we find fault with Johanson. In his original published description, he described the afarensis hand and fingers as being Homo-like,49 but once more he was seeing things hoped for, and features he had subconsciously read into the bones.
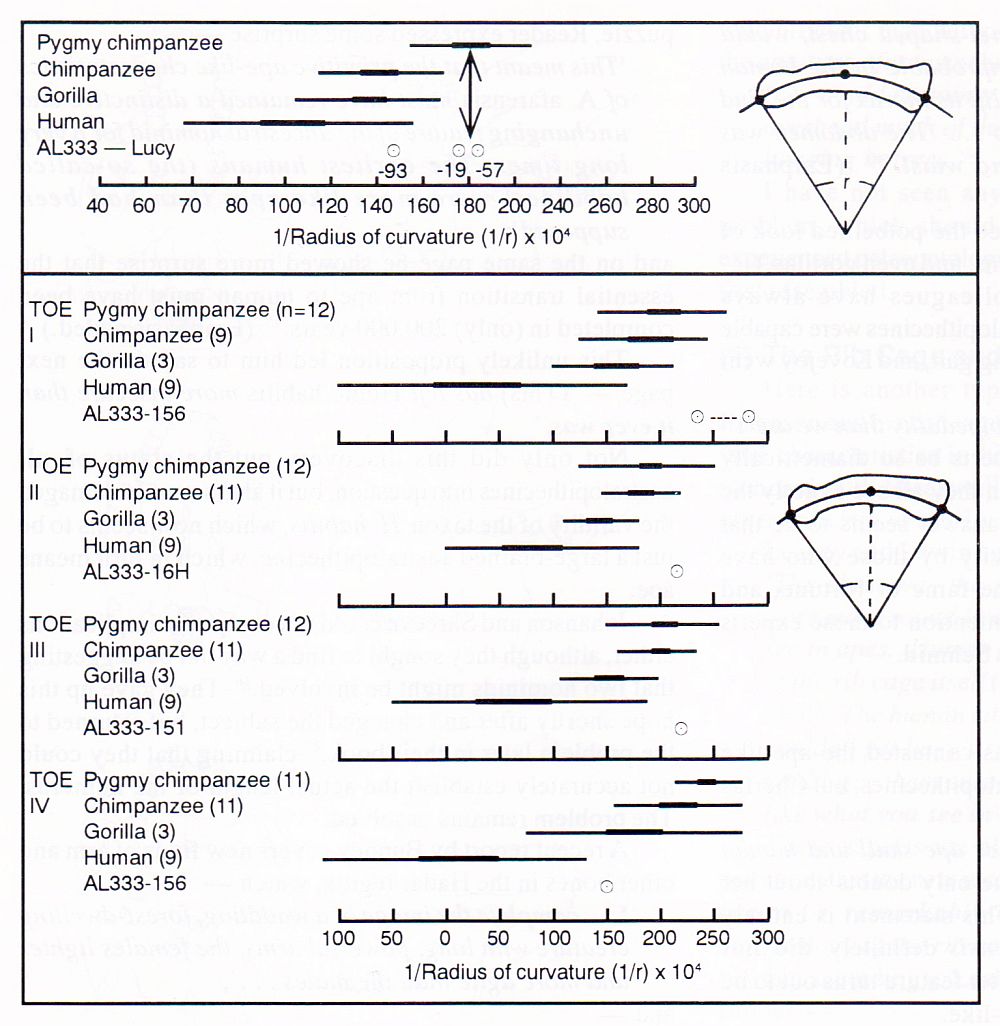
We turn again to Cherfas—“Lucy’s fingers are slender and curved … strong grasping muscles… [this] hominid is very like modern chimpanzees”. In actual fact we find the radius of curvature of the index finger lies squarely in the chimp range, and well outside the human range (see Figure 5).50
Neither did this fact escape the notice of Susman and Stern of the University of New York at Stony Brook. They concluded that Lucy and her type were more arboreal than terrestrial,51 although they did state that Lucy was close to the missing link.
Leakey and Lewin state that A. afarensis and all australopithecines were adapted to tree climbing because of at least six vital characteristics, including the fingers, waist and toes.52 Willis cited a New York anatomist: “Jack Stern offered 10 to 1 odds that afarensis was arboreal”.53
Johanson later contested these and similar conclusions by Jungers, Tuttle, Prost, Senut and Tardieu,54 and his Institute of Human Origins sponsored a scientific conference in the spring of 1983. Together with colleague Tim White, he proposed that all these ape-characteristics might be nothing more than ‘evolutionary baggage’, that is, traits left-over from a chimp-like past, and in the end he put the question in the back of his mind.55 (Neither side was convinced by the arguments of their opponents, and this ‘explanation’ will be considered again later.)
(6) The pelvis, hip and thigh
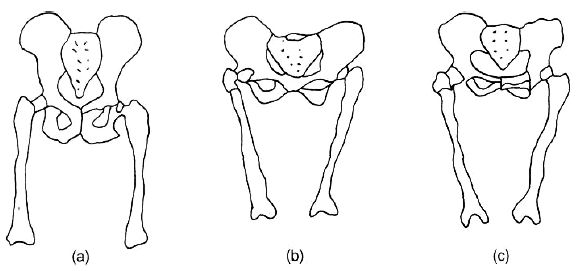
Of all the various traits of australopithecines, the pelvis is probably the most often cited by some evolutionists in support of the bipedal hypothesis regarding Lucy, yet even here we find considerable differences of opinion. (For comparisons of the australopithecine pelvis—sacrum AL288-lan and left innominate AL 288-lao—with that of a chimp and a human, see Figures 6–9.56–59

|

|

|
Johanson and his supporters often refer to the afarensis pelvis as being much more human-like than ape-like. For instance, McGowan of the University of Toronto stated:
“Australopithecines have the same cranial, femoral, and pelvic features as we do; features that are associated with erect posture.”60
Yet there are a number of evolutionist scientists who have quite important reservations about these claims. It is true that in the anterior view of the pelvis there is a certain resemblance to a human pelvis. Lucy’s pelvis is shorter and broader than that of the common chimp. However, when compared with the modern pygmy chimp, Pan paniscus, there is closer agreement.
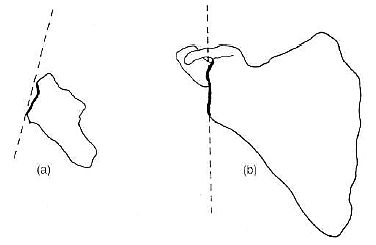
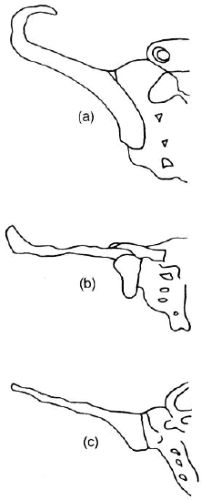
Lovejoy points out that there is a greater lateral flare of the blade than we see in humans,61 and he used this point to claim that Lucy’s alleged bipedality may have been more efficient than ours! Yet from different angles, it is clear that the afarensis pelvis is more chimp-like. For instance, the iliac blade viewed from above lies flat like a chimp, whereas in humans the blade ‘hooks’ forward (see Figure 10).62
Of the blade, Susman and Stern have this to say:
‘… the fact that the anterior portion of the iliac blade faces laterally in humans, but not in chimpanzees, is obvious. The marked resemblance of AL 288-1 (Lucy) to the chimpanzee is equally obvious [emphasis added].”63
Cherfas remarks that because the iliac blade is flat, this suggests that Lucy had a “… leaning posture like the chimp when bipedal”.64
Now a change of locomotion from the quadrupedal movement of an ape to the bipedalism of humans is ‘… no easy accomplishment. It requires a fundamental reconstruction of the anatomy, particularly of the foot and pelvis … It cannot be achieved by the “easy route” of retaining a feature already present in juvenile stages. For a baby’s legs are relatively small and weak, while bipedal posture requires enlargement and strengthening of the legs [emphasis added]”, as Gould puts it.65
My understanding of Gould means that there are no gradual stages by which such a vast change occurred—the onset of human-type bipedalism had to be sudden and huge—an absurd contention. The bipedalism of Lucy/afarensis is of the ape-type opportunistic pattern whereby many apes and even some monkeys occasionally locomote bipedally in a shuffling and uncomfortable manner for short periods.
Figures 11, 12 and 13 show the pelvis of the chimp, afarensis, and humans.66–68 Note the greater lateral flare of Lucy’s ilium.

This will become clearer as we examine the leg and foot of the australopithecines, and when all the evidence is considered, the overall picture plainly indicates their ape-type, not human-type, condition.

|

|
(7) The leg, knee and foot
As mentioned above, Susman and Stern had strong reservations about the afarensis gait. We have already seen that australopithecines had short chimp-like legs and long arms, and when we come to the knee we find that it was found 60 to 70 metres lower in the strata, and two to three kilometres away from the other parts of the skeleton,69 so we have no guarantee whatsoever that it has any connection at all with the Lucy skeleton.
Johanson and Edey provide several diagrams of the stratigraphy of the Hadar fossil area, which plainly shows that the knee joint in question was found in a stratum geologically well below that which contained Lucy’s pelvis, which also in turn was above the level of the ‘first family’ members of afarensis (see Figure 14).70
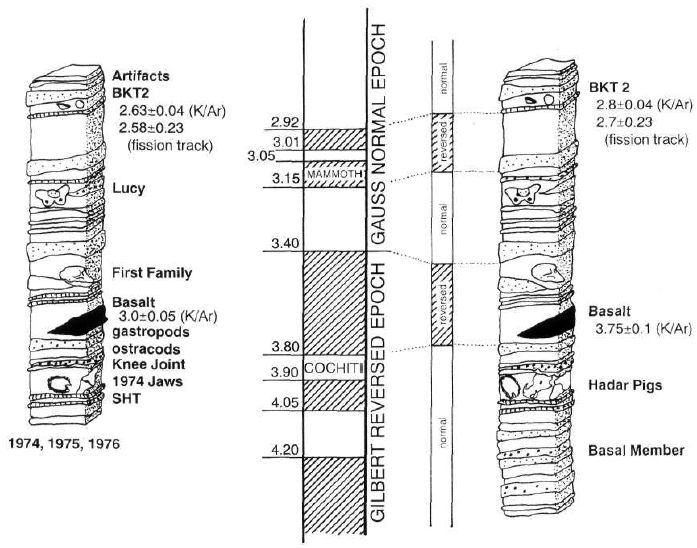
The final, corrected stratigraphic ‘column’ produced in 1979 and 1980 by palaeomagnetic and biostratigraphic methods shows large time-differences between these fossils if one accepts the modern, historical geological ‘column’ as do Johanson and Edey. They concede that:
“[This] would make Lucy and the ‘First Family’ close to 3.5 million years old; the jaws and knee joint close to 4.0 million years old [emphasis added]”.71
How then can they put the knee joint, the jaws, and Lucy’s skeleton together from sites 162, 199, 200, 333 and 400 as one single species, when there is an openly-admitted 500,000-year geochronological time interval between the various parts? These facts throw the whole assemblage of Hadar fossils into extreme doubt, to say the least!
Some Hadar ‘hominids’ (AL 162-28, AL 288-1 and AL 166-9) have strong affinities with pygmy chimpanzees, while others (AL 333-105 and AL 333-45) are more ‘gorilla-like’ in general morphology, although it is not being claimed here that modern apes are necessarily conspecific with the australopithecine specimens.
All these fossils are found in at least three separate formations—the Sidi Hakoma (mandibles and palates), and the overlying Denen Dora and Kada Hadar Formations (cranio-facial material, and the Lucy partial skeleton), which altogether cover a period of allegedly half a million years. Susman and Stern had more to say about the afarensis knee:
“… the knee of the small Hadar hominid [‘Lucy’] shares with other australopithecines a marked obliguity of the femoral shaft relative to the bicondylar plane, but in all other respects it falls either outside the range of modern human variation … or barely within it … Since, aside from the degree of valgus, the knee…possesses no modern trait to a pronounced degree, and since many of these traits may not serve to specify the precise nature of the bipedality that was practiced, we must agree with Tardieu that the overall structure of the knee is compatible with a significant degree of arboreal locomotion [emphasis added].”72
We must look, then, at the valgus degree of thigh and knee—the extent to which the leg can flex at the knee, that is, how the legs carry the body weight. In the case of the chimp this angle is zero, with the thigh and lower leg forming a straight line, with the centre of mass of the body falling inside the legs. In humans the angle of valgus is around nine to ten degrees, placing the lower leg directly under the body’s gravity centre. Lucy’s valgus angle is around 15°, which is high. Prost concluded that Lucy’s greater angle favours other supporting evidence that Lucy and other australopithecines were adept tree climbers.73
We should remember that the cited valgus angle for afarensis is based on measurements of the fragment AL 129-a, which is questionable because of the distance separating it from site 288. Another femoral fragment from nearby site 333 (the ‘First Family’), and which is included in the A. afarensis composite, yields an angle of only 9°.
Prost pointed out that among primates, the spider monkey and the orang have about the same degree of valgus as humans; yet both are extremely agile in the trees. It seems the valgus angle means little. Prost, Susman and Stern nevertheless argue that Lucy’s angle reflects her ability to climb, and that her bipedal gait, when she used it, was very much like that of a chimp or spider monkey.74 For much the same reasons Tardieu also believes that all the smaller afarensis specimens must have spent a lot of time in the trees, like orangs and monkeys.75
Susman recently observed that skepticism is warranted, even if the knee did belong with the rest of the Lucy skeleton, because Lucy’s proportional foot length was 30 per cent greater than that of the average human: “[this means] greater bending at hip and knee joints, resulting in a high-stepping gait … Lucy lifted her legs rather awkwardly while walking—like a modern human wearing a pair of flippers”,76 and this fits remarkably well with the other views cited above—Lucy locomoted on the ground like a chimp. Because the fingers and toes were curved, with marks of strong grasping muscles and large tendons, Cherfas comments thus:
“In these respects, the hominid (A. afarensis) is very like modern chimps … Stern and Susman conclude that while she probably used her hooklike hands and strong wrists to climb in trees, she did not knucklewalk on the ground [emphasis added].”77
Whether she ‘knuckle-walked’ or not is debatable; although Johanson et al. do not accept that she did (this would be too much of a chimp-like feature for them to accept), Shea is not so sure. The traits concerned may not always show up in fossil material— the matter is inconclusive.78
(8) The ankle (talus)
This is another vital piece of evidence to be considered; the joint of the tibia and the foot. It appears that the small afarensis specimen had a very ape-like talus. In the larger specimens the talus has a forward tilt, while Lucy’s ankle tilts backwards even further than a gorilla’s. In humans the joint faces slightly forward for ease of an erect full-striding gait.79
While it is true that a backward-angled talus does not prevent a forward bending of the foot, it would cause some leaning over and awkwardness, chimp fashion, with bent knee and hip while locomoting erect. With such a talus there is no way Lucy could stride out in human fashion. The matter of the forward-facing tilt of the larger afarensis specimens is frankly a mystery, but there is simply no way by which the larger, supposedly male specimens would be so much more advanced than the smaller female specimens. Male and female individuals of the same species could hardly have evolved at such a disparate rate, and I was surprised that Cherfas, in a personal communication to me, would put forward such an idea.80
Of the foot of OH 8, a habiline, Oxnard and Lisowski say that the Olduvai foot is not adapted for bipedality in the manner of man, and that it displays features (which resemble) the feet of arboreal creatures, and that when it walked bipedally it locomoted with flattened arches rather than with the high arches of man.81 This conclusion is strongly supported by Wood—the foot of OH 8 (and OH 10 and OH 35, all fragments of one individual) is very ape-like.82
The Laetoli Footprints
In 1976, Mary Leakey uncovered a trail of human-like footprints at Laetoli, about 30 miles (48 km) south of Olduvai,83 and dated at well over 3 Ma. Of these prints, White says:
“Make no mistake about it … they are like modern footprints. If one were left in the sand of a Californian beach today … [one] would instantly say that somebody had walked there … [one] wouldn’t be able to tell it from a hundred other prints on the beach [emphasis added]”.84
Tuttle of the University of Chicago saw an “unresolvable discrepancy” between these prints and the foot of Lucy/afarensis, who, most experts believe, made them. Tuttle concluded that the prints had been made by a creature walking like humans, and therefore that a ‘true Homo’ had existed at Laetoli, and left its tracks behind.85 Although White disagrees with this,86 Tuttle, an expert in this field, insists he is right—“In sum, the 3.5 million year old footprint trails … resemble those of habitually unshod modern humans [emphasis added]”.87
In a later article his comment is as follows:
“In discernible features, the Laetoli prints are indistinguishable from those of habitually barefoot Homo sapiens.”88
A year or so later, Tuttle and Webb reaffirmed his previous statements:
“[Casts] … further illustrate the remarkable humanness of Laetoli hominid feet in all detectable morphological features [emphasis added]”.89
According to Lubenow, a creationist, Tuttle made two other attacks on the prints.90–91 Tuttle’s verdict is:
“If the [Laetoli] G footprints were [not] so old, we would readily conclude that they were made by a member of our genus, Homo [emphasis added]”.
There we have it! Because of their supposed age of 3.7 Ma, the evolutionist simply cannot accept them as human—mankind is not supposed to have existed then, despite the conclusion that the known hominids such as Lucy/ afarensis didn’t make them. Such is their faith in evolutionary theory and in evolutionary geochronology, that they cannot accept what is staring them in the face. Once we rid ourselves of the mental shackles of million-year-old geology, there is nothing to prevent us from taking these footprints at face value. That is the only way to resolve the problem, and the view that full humans did make them ties in rather well with the fully human femurs found in Java (1.8 Ma) and at Koobi Fora (2 Ma).
Although Oxnard has been attacked by fellow evolutionists, the fact remains that his morphometric analysis of australopithecine tali, bones vital in locomotion, indicated a gulf between modern man, the habilines, the australopithecines and modern African apes. In fact, australopithecines had tali most like those of the tree-dwelling ape, the orang-utan. There was no indication that the australopithecines were intermediate between humans and apes.92
This view is still held by Oxnard, who in 1984 said that:
“… the australopithecines (from Olduvai and Sterkfontein, Kromdraai and Makapansgat), are now irrevocably removed from a place in the evolution of human bipedalism, possibly from a place in a group any closer to humans than to African apes, and certainly from any place in the direct human lineage [emphasis added].”93
In the same book, he considers that some of the australopithecines form a radiation separate from either humans or apes—they are not structurally closely similar to humans, they must have been living, at least in part, in arboreal environments.94 By 1984 Oxnard was well aware of the Lucy/afarensis fossils and was writing about studies of those specimens. He is most certainly not a creationist and still believes in evolution, but he finds no human ancestry in the australopithecine group. In view of the evidence assembled here, one must take Oxnard’s hypothesis seriously—that these peculiar creatures were not intermediate between ape and man, but were extinct, previously unknown pongids.
| Name | Brain Capacity (where known) | Fossil Material |
|---|---|---|
| OH 24 | 590 cc | cranial |
| OH 7 | 670 cc | parietals, mandibular, post-cranial |
| post cranial OH 13 | 670 cc | cranial, mandibular |
| ER 1590 | 750 cc (?) | cranial |
| ER1805 | 510 cc | cranial |
| Stw 53 | — | skull fragments, teeth |
| (ER 1470) | 750 cc (?) | skull |
| (OH 8) | — | post-cranial |
| (OH 62) | — | skull and post-cranial fragments, limb bones |
Table 2. Some of the most important habiline fossils. The evolutionary time-scale is 1.8–2.0 million years. Questions remain about the cranial capacity of ER 1470 and possibly ER 1590 because of a recent skull reconstruction of ER 1470. Skull ER 1590 (similar to ER 1470) possessed a saggital keel as do most male apes. See text for discussion.
The problematic habilines
Since the so-called habiline specimens KNM-ER 1470, OH 8 and OH 62 have already been referred to, the discussion here will be restricted to six other ‘best-preserved’ specimens, as set out in Table 2.
The question is—are these remains, which are generally referred to H. habilis, true intermediates between the australopithecines and Homo erectus, and therefore human ancestors, or are they simply larger-brained australopithecines? Are they composites of true H. erectus forms and australopithecine apes? Or again, did many of the authorities ‘see’ human-like features which were not really there?
Of these questions, Wood says:
“It is remarkable that the taxonomy and phylogenetic relationships of the earliest-known representatives of our own genus Homo, remain obscure … [a simple unilinear] model of human evolution, in which Homo habilis succeeded the australopithecines and then evolved via H. erectus into H. sapiens [is] untenable—but no clear alternative consensus has yet emerged”.95
Even in the early stages this taxon attracted criticism and controversy, with some authorities believing that H. habilis exhibited “… too few advanced features to separate it from Australopithecus”;96 while “… others complained that some of the [specimens] were indistinguishable from H. erectus”.97
On the same page Wood asks of these specimens: “Do they belong to Homo, or is one or more an australopithecine?” On the next page Wood again asks: “H. habilis—one species or two?”98
Controversy and disagreement continued to rage among the experts through the 1970s and 1980s in respect of the taxonomic unity of the group, with some arguing that H. habilis should be restricted to OH 7 and OH 16, and that others such as OH 13 and OH 24, ER 1805 and ER 1813 were “… judged to be late-surviving small Australopithecus individuals that were contemporary… with H. habilis [and] with H. erectus”.99
Wood himself argues that the type specimen resembles, though is not necessarily conspecific with, H. erectus, and puts forward the name Homo ergaster as the proper name for the probable African precursor, (or variety of) H. erectus; a taxon which would also include the crania ER 3733, 3883, and probably the skeleton KNM-WT 15,000 (a true H. erectus specimen);100 that is, the habiline type specimen is really H. erectus, as is ER 3733.
On the next page Wood points out that some of the Koobi Fora specimens’ leg bones exhibit “… later Homo-like morphology (which) contrasts with that of the more australopithecine-like morphology of the Olduvai remains.” (Certain ‘derived’ features appeared in some of the Koobi Fora remains which were found alongside KNM-ER 3735, which itself resembles the primitive ape-like OH 62 skeleton.) The two Koobi Fora femora are like modern humans and archaic H. sapiens, and are very different from predictions based on the australopithecine-like Olduvai habiline finds.
This, Wood says, points to taxonomic heterogeneity in ‘early’ Homo, and on the next page,101 it becomes clear that because of wide morphological variability, Wood favours breaking up the Olduvai and Koobi Fora members of the taxon into A. africanus, H. erectus, H. habilis (Group 1 large specimens); H. habilis (Group 2 small specimens); H. rudolfensis, H. ergaster, H. habilis (sensu stricto), and Homo sp.
One of the problems seems to be that originally H. habilis was defined in terms only of what was known at the time—that is, australopithecines, H. erectus (Java and Peking), and H. sapiens. In fact, Louis Leakey ‘created’ H. habilis by changing the definition of Homo to suit his find.102 Even today there are few explicit definitions of the Homo clade.103 According to Wood, an early (1975) study suggested that an ECV of more than 600 cc was sufficient to distinguish H. habilis from A. africanus, while other experts were unable to identify any suitable features because no distinction was then being made between gracile australopithecines from southern Africa and the Olduvai habilis specimens.104 Also see Johanson and Edey.105
More recently however, a narrow mid-face, an elongated anterior basicranium, and a mesiodistally elongated crown in upper P1 have been put forward as distinguishing features. Wood offers a cladistically-defined set of eight characteristics for distinguishing Homo, and five more for a sister group, which a hypothetical common ancestor for H. habilis (sensu stricto) and H. rudolfensis, the two subtypes he asserts make up H. habilis, would share.106
On the same page Wood maintains that if the taxonomic integrity of ‘habilis’ is to be retained, then “… it is a species that manifestly embraces an unusually large amount of variation. … Those who believe this range to be unacceptably wide, and thus for whom H. habilis represents more than one species, disagree about how the hypodigm should be apportioned [emphasis added].” (The hypodigm is made up of the various specimens allocated to the taxon.)
The evolutionary time-scales involved range around 1.9 Ma, which is little different from the ‘earliest’-known evidence for H. erectus, or H. ergaster, which is the species name Wood applies to some of the material others call erectus. (Wood believes ergaster was an earlier type.) Note also that recent finds in Java indicate that H. erectus/ergaster already was in that region as long ago as 1.8 Ma.107
Wood suggests that H. habilis (sensu stricto), H. rudolfensis and H. ergaster (erectus equivalent) have evolved separately from different australopithecine ancestors, thus giving us no less than three Homo lines in the Upper Pliocene and Lower Pleistocene systems!
In view of all this confusion, and in view of the fact that a clearly human ‘erectus’ existed at least from around 1.9 Ma in Africa (to say nothing of Java), and considering the ape-like primitiveness of habilis OH 62 at 1.8 Ma, giving virtually no time to change to man, could we not equally say that individuals classified by some as habilis, with a large range of variation, may represent contemporaneous populations of both Homo erectus (human) and australopithecines? This would mean that ‘habilis’ is a phantom, as Wood more or less unknowingly concedes when he repeatedly refers to the idea of habilis being more than one species.
Harris has apparently become disillusioned:
“Habilis turns out to have been only a little over three feet tall—just like the diminutive afarensis named Lucy. And it still had somewhat curved toes and fingers, long arms, and short legs. … Except for its bigger brain and its association with stone tools, habilis is virtually indistinguishable from the earliest australopithecines. This raises doubts about whether habilis should be regarded as a member of the genus Homo [emphasis added].”108
The statement regarding tools is only an opinion expressed by Leakey.109 The tools could have been associated with other types (perhaps with true humans who may have made the Laetoli footprint trail?).
If ER 1470 was only a large-brained australopithecine with an exceedingly ape-like face as prognathous as Lucy’s (and therefore so were the other later specimens), and dating from 1.8 to 1.9 Ma, then how strong is the case for other habiline specimens to be ancestral to Homo erectus? Because one large habiline specimen (ER 1470) and one of the smaller ones (OH 62) have now proved to be so apelike (like Lucy and others), what grounds remain for the other specimens to be classified as Homo? These two specimens are among the most important in the habilis taxon!
It is clear that Richard Leakey should have been much more cautious in his original reconstruction of 1470, because at the time (1972) he was already aware of the dangers of preconception, and he acknowledged that the angle of face-to-cranium was uncertain. His colleague Walker remembers:
“You could hold the maxilla [upper jaw] forward and give it a long face, or you could tuck it in, making the face short.… how you held it one way, it looked like one thing, if you held it another [way] it looked like something else.”110
We had to wait until 1992 for its true appearance to be revealed by Bromage. Walker himself of course has always insisted that 1470 was too much like Australopithecus to be ignored. (The limb bones associated with 1470 were found a considerable distance away from the skull itself, and thus we cannot definitely establish whether they belonged with the skull or not.111)
Leakey once said that some of the smaller habilines (including ER 1813 and possibly OH 24) could be justifiably downgraded to gracile australopithecines,112 while Walker is satisfied that ER 1813, OH 13 and OH 24 are only late-surviving forms of gracile australopithecines.113 Brace, Wolpoff and Montagu are all agreed that ER 1470 is only a large-brained australopithecine—A. africanus.114–115
Stringer has broken up the habilines into two groups—the large, ‘habilis-like’ specimens ER 1470, ER 1590, ER 3732, OH 7 and OH 24, and the smaller types ER 1805, ER1813, OH 13 and OH 16, which are more australopithecine-like.116
Whatever else may be said of the enigmatic specimen ER 1813, its post-cranial features are very similar to those of OH 62. Whatever the various authorities choose to call these creatures, australopithecines or habilines, the fact remains that apart from brain-size in a few of them, they all still overwhelmingly display the same chimp-like features of the ‘early’ specimens.117 Further to be considered is the fact that both 1590 and 1805 possessed saggital crests, which are strong pointers to ape-type robustness.118 Beasley believes that habilis and some of the graciles may be conspecific, such as 1470, 1813, and Sts 5.119
We simply do not have enough post-cranial material for 1470 to say with certainty one way or the other, but going by its reconstructed skull, the odds are strong that it too is an extinct ape, and now that we know that the larger specimens are very ape-like anyway, what does this mean for the smaller specimens? It seems the whole taxon should be scrapped!
The recent news that H. erectus was already in Java just under 2 million years ago makes the whole proposition a farce, and Lewin says that the current theories are “in ruins” as a result of new dating of Java Man fossils.120 See also Holloway.121
Either the evolutionary time-scales are completely wrong and useless, or the guesses about mankind’s origins are no more than speculation and preconceptions, or both. It seems the discontinuity between man and the apes is still unbridged, despite all the hype.
Somewhat surprisingly, Groves believes that the larger-brained habiline of the ER 1470 type was not closer to the human line. Groves plumps for the smaller-brained ER 1813 type.122 On the same page he tells us that there were three species of habilis—the ER 1470 type, the ER 1813 type, and the Olduvai specimen OH 24!
Have all the habilines been over-rated, like 1470 was, before Bromage found its true morphology? Have the palaeontologists and palaeoanthropologists been seeing Homo affinities which were not really there, as in the case of Piltdown ‘Man’ before it was found to be a hoax, and with Zinjanthropus, Lucy and many others? Was the remarkably modern femur found in the same general region as ER 1470 really a human thigh-bone, rather than belonging to some ape-like creature?
All these questions remain unanswered at present, but the case for Homo habilis, as we have seen previously, must be in serious doubt, as admitted by Reader.123
New findings
Another key piece of evidence to be considered comes from a recent study of hominid (inner) ear morphology by a team led by Spoor, a Dutch anatomist of the University College, London, and reported in a 1994 issue of New Scientist.124 After reporting that there is still no consensus on the matter of bipedalism in the hominids, Shipman goes on to mention the results of a long examination of the bony labyrinth of the inner ear of hominids and humans. Spoor’s team concentrated on the part of the vestibular system known as the semicircular canals—three bony tubes which curve through the bone that underlies the external ear.
These canals have a lot to do with balance while locomoting erect. Spoor and his team, using high-resolution computerised tomography, scanned a large number of specimens, including A. africanus and A. robustus, plus two specimens of habilines in addition to a H. erectus specimen, as well as other primates from small squirrel monkeys to gorillas—over 100 specimens of extant or extinct primates of known locomotor habits. According to Shipman’s article, the scans of all australopithecines and habilines told a consistent story—that the bony labyrinths were decidedly ape-like, but in contrast, the canals of H. erectus were identical to those of modern humans. None were ‘intermediate’. Spoor’s team believes that the australopithecines might have balanced on two legs when standing, rather than when moving, just as chimps do when gathering food.
The most interesting result was from the scan of a H. habilis specimen, Stw 53 from Sterkfontein. Spoor says:
“It’s very difficult to interpret; the only thing that the labyrinth suggests is that [H. habilis] is less bipedally adapted than the australopithecines. It looks much more like gibbons, maybe, or like baboons, certainly not a human pattern [emphasis added].”125
On the same page Shipman says:
“[Spoor’s] conclusion is that either this specimen is not H. habilis, or if it is, H. habilis is unlikely to be ancestral to [humans] [emphasis added]’. (Emphasis added.)
According to another report in Science News:
“Australopithecines are more similar to chimpanzees than to modern humans in their inner-ear morphology … Inner ears like those of modern humans first emerged in Homo erectus [emphasis added]”.126
I have recently argued that so-called H. erectus is so close morphologically to modern humans that it should be included in the same species as H. sapiens, and these findings confirm that claim.127 No other hominid possessed the human-like canal proportions, thus tending to confirm that neither the australopithecines nor the so-called habilines have anything whatsoever to do with the origin of human beings.
In Shipman’s article referred to above, anatomists Martin and Hartwig-Scherer of Zurich University, welcomed the Spoor findings which strongly support their analysis of habiline limb proportions. Martin says:
“The combined evidence now suggests that we in fact have before us the remains of a distinctive hominoid from Africa, but of a great ape rather than a hominid.”128
Thus we now have clear evidence to support the strong case against any australopithecine being the evolutionary ancestor of humans.
My conclusions are in close agreement with Zihlman’s analysis of the afarensis skeleton—she also found remarkable similarities between the skeletons of the pygmy chimp and Lucy. The only questionable differences are found in (some) parts of the pelvis, and in a couple of the dental features—much too few on which to base an evolutionary case.
General comments
It is realised of course that evolutionary theory would predict the existence of a creature which showed a mixture of human and ape characters—an ideal ‘intermediate’, so to speak, and Johanson and others have presented Lucy/afarensis and other australopithecines as just such an intermediate type. The further we go back time-wise, the more ape-like and less human4ike such a ‘common ancestor’ should become.
Unfortunately for the theory there is a serious flaw in this argument, in addition to the strong doubts we have already seen above. If bipedalism was established by 3.3 Ma (A. afarensis), then it follows that her descendants should have become more and more human-like in their postcranial morphology, and had fewer and fewer pongid features as time passed. We recall the claim by Johanson and Shreeve,129 that certain ape-like features of the afarensis specimens, such as the long, curved digits, the ape-like shoulder and other simian features, were only ‘evolutionary baggage’—leftovers of a chimp-like past. The trouble is that the so-called ‘evolutionary baggage’ is still there, even in the most recent australopithecine and ‘habiline’ specimens.
There has been no progression towards humanness in any of the lines—africanus, robustus, boisei or the hyper-robust specimen commonly known as the Black Skull (WT 17000). In fact, between the afarensis finds dated at over 3 Ma and the habiline OH 62 at about 1.9 Ma, we find only evolutionary stasis! Africanus is believed to have become extinct around 2 Ma, robustus/boisei about 1 Ma; and the only hyper-robust specimen WT 17000 is believed to date from about 2.5 Ma.130
One would have expected that if australopithecines were intermediate between humans and African great apes, these animals would fill or substantially reduce the large morphological and behavioural gap which separates humankind from the pongids. However, even in the best possible light they don’t come close. Only in the pelvis and in brain size is there a possible argument, and even there the case is marginal at best. The larger brain capacity of one or two of the ‘ habiline’ specimens (around 680–750 cc) is nothing to get excited about if modern gorillas can range from about 450 cc to 750 cc. If man’s capacity covers the range from about 900 cc to well over 2000 cc, then we should not be surprised if a few australopithecine individuals also show considerable variation. Montagu and Brace were not fazed—they declared this habiline specimen (ER 1470) to be nothing more than a large-brained australopithecine,131 while Clark takes a somewhat similar view.132 There are just too many overwhelmingly simian features in all these curious animals.
The more rounded skull of some africanus specimens is probably nothing more than a fortuitous case of negative allometry, and the prognathicity of these creatures is far more ape-like than human-like—in fact, their faces and jaws thrust forward even more than the robust specimens. Of the afarensis species Kimbel says that: “There is no obvious sign of evolution in this prehuman species for about 1 million years, yet later, in only a fraction of that time, A. afarensis gave rise to a great branching of the family tree.”133 Kimbel was referring to new afarensis finds, and the ‘great branching’ referred to was the other australopithecines (africanus, robustus and boisei).
This is confirmed by Lewin, who points out that “A number of researchers consider that the Laetoli/ Hadar fossils are very similar to, if not identical with, fossils from one of the South African sites”, even though a million years of evolution supposedly took place.134
Eldredge recently commented on the dentition of the gracile africanus:
“The massive build of the face and the huge teeth bespeak something more along the lines of a gorilla than a human”.135
Needless to say, we find the robustus features are just as ape-like. In fact, the differences between the various australopithecine species are in most cases so slight that one could lump them all together in one group, except possibly for brain size. The largest ECV recorded for any alleged prehuman is that of skull ER 1470 with a capacity of about 752 cc, compared with Lucy’s estimated size of about 450 cc. In view of Bromage’s reconstruction of ER 1470 (see later), even 752 cc may now be too large an estimate.
It is commonly known that a modern gorilla was recorded at 750 cc, so skull ER 1470, an alleged habiline, had a brain size no bigger than an extant gorilla, and in view of Bromage’s recent reconstruction of this skull exhibiting so many pongid features, why class it as a ‘Homo’ at all?
Even the allegedly most Homo-like specimens, the habiline ER 1470 and others of around 1.9 Ma, have the same evolutionary ‘baggage’, especially in the light of Bromage’s recent work. Johanson himself says of the habilines that they would be so repulsive in looks that if one boarded a suburban train, people would move to the other end of the carriage.136 Surely this is a very appropriate comment, and yet these ape-like creatures are supposed to be the most advanced of the alleged human ancestors! Imagine then what the older australopithecines must have looked like!
The real puzzle in all this is that, apart from minor variability in the genus Australopithecus with its three or four species, there is no sign of any of them evolving into human beings. Bunney is intrigued by these problems and she wrote recently:
“The puzzle is how the form represented in the hominid OH 62 (a habiline) and its male equivalent … could have evolved in 200,000 years or less, into a hominid as large as Homo erectus”.137
The reason for such frankness has to do with the discovery of the Turkana ‘Boy’, by Richard Leakey in 1984, and classified as Homo erectus. This specimen, called WT 15000, is clearly human and is remarkably complete. Though of a young individual, he may have been over six feet tall when fully grown.
Although the various types of australopithecine do exhibit relatively minor cranial and post-cranial variability, it is quite impossible to trace any sequential ‘evolution’ towards humanity. It was quite a setback therefore when it was found that OH 62 possessed bodily features just as primitive as Lucy/afarensis, which lived allegedly well over one million years before, and therefore it seems that the so-called habilines are no more than australopithecine varieties.
In the case of the ER 1470 skull, the original reconstruction by Richard Leakey had a slightly higher forehead and a flattish face, but even then, creationist Hummer was cautious about this find. As long ago as 1977 he pointed out a number of australopithecine features—the long ape-like upper lip; the cranio/facial index of 59.0 which is in the middle of the australopithecine range (51.0 to 64.5), while in humans the index range is 30.0 to 45.0; and the skirt-like profile of the skull so characteristic of australopithecines. In fact, the general similarity to A. africanus is striking.138
Brace firmly believes that habilis and A. africanus are conspecific—that is, they are one and the same species.139
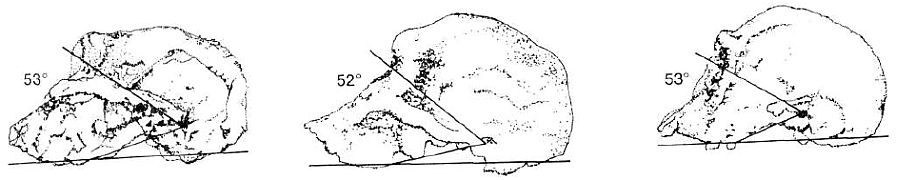
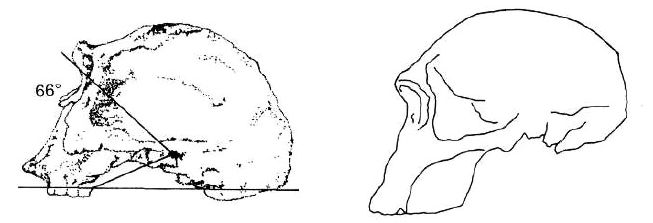
In the new reconstruction by Bromage already mentioned, a new procedure was used which shows that Leakey’s original casting was totally incorrect. Bromage is an expert in the study of bone growth processes. His work indicates that the so-called early Homo-type material (ER 1470, the habilines and A. africanus) had facial development patterns typical of monkeys and apes. Using computerised scanning techniques, Bromage made the new reconstruction, and the result is that these specimens were very apelike and all look like typical australopithecines (see Figure 15). This leaves evolution without a connection between Homo and the apes.140
| Meatus Angle | Approximate Brain Capacity | |
|---|---|---|
| Chimp | 49° | 400 cc |
| A. afarensis | 46° | 350 cc |
| A. africanus—(Sts5) | 47° | 450 cc |
| A. aethopicus—WT 17000 | 46° | 350 cc |
| A. africanus—(Sts71) | 53° | 400 cc |
| A. boisei | 53° | 455 cc |
| H. habilis—(ER 1470) | 52° | 750 cc (?) |
| H. habilis—(ER 1813) | 53° | 750 cc (?) |
| Homo erectus—(a human) | 66° | 900–1250 cc |
Table 3. The results of Bromage’s research indicates the close relationship of apes, australopithecines and so-called habilines. The ‘meatus’ angles (pitch of the face to the cranium) for all apes, australopithecines and habilines lie within the range of 46–53°. Humans are clearly different, with even the allegedly ‘primitive’ H. erectus displaying 66°. Questions remain over the figures for ER 1470 and ER 1813 because of the reconstruction of the former which could now yield lower figures for brain capacity. This could also apply to ER 1813, making the distinction between the habilines and humans even sharper.
But there is even more—Bromage also analysed the feature known as the ‘meatus angle’, that is, the pitch of the face onto the cranium. There were no surprises for creationists when he discovered that in all australopithecines, including so-called habiline specimens ER 1470 and ER 1813, the meatus angle is very close to the chimpanzee figure, and a more human angle is not reached until the Homo erectus specimen ER 3733! The figures are as set out in Table 3.
Bromage concludes that “… the development of early hominids must have been more like apes than modern humans.”141 To be sure, Bromage is not abandoning evolution—on the next page he says that ER 1470 is Homo in many respects, and it has a phenomenally large brain for its time, yet skull 1470 and Taung are now so chimp-like (as he concedes)142 that it surely must be increasingly difficult for the palaeoanthropologists to keep maintaining their position. It appears they are so paralysed by the concept of evolution that they cannot think any other way. Most of them have to ignore their very own analyses and computations!
A highly significant example of just such a case is that described by creationist Lubenow. The anatomist and anthropologist Lovejoy openly admitted that he had disregarded his very own multivariate analysis of Lucy’s knee-joint because it showed that it was far-removed from man, and was right in the middle of the ape range.143 This plainly demonstrates my point—Lovejoy had obviously expected the computer analysis to indicate it was modern and human-like, and he could not accept that it wasn’t.
The origin of humans
Far from establishing australopithecines as being our ape/man evolutionary ancestors, I believe the overwhelming evidence from these fossils points in the direction of the smaller specimens being close to the ancestors of present-day pygmy chimpanzees, Pan paniscus, with some of the coarse specimens of A. robustus being the forerunners of, or close relatives to the common chimp, Pan troglodytes, and the larger, very coarse forms such as A. bolsei being closely related to modern gorillas.
At least, given the usual wide range of genetic and morphological variability within genera, these three extinct ape-like creatures could be very close relatives of these three modern great apes. The plain fact is that we otherwise have no suitable ancestral forms for modern apes, while at the same time, we have no suitable descendants for the three australopithecines. This would leave man as always man, and ape as always ape. The australopithecines have all gone extinct apparently without issue, and we have no ancestral chimpanzee or gorilla fossils!
In view of the repeated admissions of the chimp-like or gorilla-like characteristics of all australopithecines, British experts Cherfas and Gribbin were forced to the conclusion that these extinct forms were the very close historical relatives of today’s gorilla and chimpanzees.144 On these pages they say:
“… no fossils of ancestors to the modern chimp and gorilla have been found; so we have two living species, without known ancestors, and two fossil species, without known descendants, and all four are closely related to man … as Holmes said, once you have eliminated the impossible, what remains, however improbable, is the truth.”
Except for the predictable comment that all are related to man, I would agree that such a hypothesis is possible, because of the considerable variability within genera and families. Beasley has argued for a biblically-based concept that there was a morphological shrinkage of various forms following the Noachic Flood, and that today’s great apes are very closely related to the gracile and robust australopithecines which were their ancestors.145
Another very real possibility is provided by a study of Oxnard’s work wherein he concludes that the Olduvai and Sterkfontein australopithecine fossil material, when compared with humans and African apes, is far more different from the equivalent parts of humans and apes, than are these from each other:
“… the fossils are indeed uniquely different from these extant hominoids. It is in this sense that I use the term ‘unique’ … This uniqueness leads inexorably to the possibility that these fossils [the australopithecines] are not ancestral to either humans or apes [emphasis added]”.146
Just eight pages later, Oxnard says that the results of the multivariate morphometric studies of australopithecines show the various specimens are more similar to each other than to any individual living primate, and further, the differences from any living form are greater than the differences between bipedal humans and the terrestrial apes.
As we have already seen a mountain of evidence that indicates the closeness of all australopithecines to modern chimps and gorillas, why not assign them all to the same non-evolutionary ancestral family as modern pongids rather than to the human family? The evidence already given here is surely sufficient to consider the strong probability that there were at least two other pongids in the Pliocene/ Pleistocene which, being very ape-like in appearance, were close relatives, but not necessarily conspecific with the African great apes. We know that none of the australopithecine species were actually today’s chimps or gorillas because of the morphology of the pelvis and some of the dentition. They may indeed have been members of a different genus which has gone extinct, as have so many other apes and monkeys of the past.
Although this is the most sensible view, based on all the latest evidence, it is unlikely to be accepted by the evolutionary establishment because it would mean that human ancestry is a mystery, and that would be a severe blow to the neo-Darwinian paradigm.
Once we acknowledge that scientists are as fallible and as prejudiced as anyone else, as is often admitted by some of the more frank researchers such as Lewin, we can see why it is that there is so much strong disagreement and controversy about fossils—the finders and their supporters naturally want their discoveries to be very important—it brings fame, fortune, and big research grants. On the other hand, those more independent scientists who have no particular axe to grind are in a better position to assess the meaning of such finds, and this is why the discoverers interpret their finds in a far different manner from those who have a clear head and a more independent attitude.
Not infrequently, and particularly in supposed human evolution, there is a tendency for the more popular science writers to gloss over or ignore the objections and problems raised by the more objective authorities, thus giving the public the impression that the whole case is proved and no real problem exists. Despite the lack of known ancestors for the African great apes and for the australopithecines, and the absence of an ancestor for Homo erectus, which seems to appear virtually full-blown from nowhere in the Upper Pleistocene system, human evolution from australopithecines has now become dogma and creationists must constantly be on the attack to remind students of the many weaknesses of the theory, and to highlight the contradictions, the speculation, and obvious bias and preconceptions, displayed by its adherents. Evolutionist Lewin’s 1989 book is packed with such examples, and it appears that just about every palaeontologist has been guilty of subjectivity—Louis and Richard Leakey, David Pilbeam, Elwyn Simons, Don Johanson are just a few.
Some late finds
There have recently been other news reports of further fossil finds from about 4.4 Ma. However, it is presumptuous to claim these finds yet again as the ‘missing link’. Since World War II, we have been told the missing link has been found so often that we have become rather cynical. It often takes several years for the original hype to give way to a more sober assessment of the finds. In fact, this has happened with the fossils found in 1992-1993 in Ethiopia, and named Australopithecus ramidus.147
The finds included fragments such as teeth, arm bones, and parts of the skull and jaws belonging to an estimated 17 individuals. The new species shares a wide array of traits with A. afarensis (Lucy, etc.), including canine teeth similar to ancestral apes and an unmistakable chimp-like skull. Wood described the finds thus:
“… [ramidus] lies extremely close to the divergence between the lineages leading to the African apes and modern humans”.148
Yet only a year later, following more finds in Ethiopia, ramidus has been renamed, reclassified in a different genus Ardipithecus, and removed from the human line!149
White now believes an even more recent find of 21 bone specimens (4.2 Ma), and which are “far more apelike than Lucy”, is now the link between apes and humans.150 (Emphasis added.) This latest find has been named Australopithecus anamensis, but the only clues to any hominid affinity are a piece of shinbone thicker than a modern human’s, and broader molars with thick enamel, while the jaw resembles that of a chimpanzee. Furthermore, in the Nature article of November 3, 1994, we find that Homo habilis has also been removed from the human lineage.151
In both of the above cases the material is not large, and it is difficult to draw any sound conclusions from it. One would think by now that the palaeontologists and anthropologists would have learned their lesson from previous experience and would refrain from passing judgment on fossil scraps too few to be making such wild pronouncements, but it appears that in their eagerness to prove human relationships with apes they have failed to learn that lesson. I have little doubt that after the passage of a few more years anamensis will also be sidelined, only to be replaced by some other dubious fossil find.
Evolution and creation
There is no scientific way to prove divine creation of genera and families of living things, just as fossils cannot be used to prove evolution, as Ridley admitted.152 However, the huge and acknowledged number of missing links between classes, orders, and families, points strongly to creation, not evolution.
The published literature of ‘human evolution’ studies reveals there is many a scientist who has not obeyed the dictum of science—to hold a theory with a light hand and be prepared to let it go when the evidence requires it. Among the names listed above are several who have doggedly held on to their pet interpretation long after the evidence made it clear that they were wrong, and they had to be dragged kicking and screaming to the correct conclusion.
Although I have referred here to uniformitarian dating for convenience, it should be noted that this does not signify my belief in such timespans; in fact I believe, on good grounds, that the earth, the Solar System and the Universe are relatively young and were created by God in the recent past; but in any case time is irrelevant in respect of the fossils—whether thousands of years or millions of years, the fossil record fails to support an evolutionary origin for human beings, and it fits the creationist case far better.

Table 4. Probable phytogenies of the African anthropoid ape family and the human family, set out in the evolutionary time-scale for convenience only. There are no established evolutionary precursors for either the African apes or for humans. Note that afarensis may have given rise to three subspecies, but it should be remembered that all australopithecines belong to a single family/holobaramin, and the morphological differences between them (and between all humans) is slight. Variability within the type does not necessarily depend on the passage of time.
| Feature | Status | Ref. | Feature | Status | Ref. | ||
|---|---|---|---|---|---|---|---|
| 1 | Foramen magnum | b. | 153, 154 | 19 | Rib-cage | a. | 163 |
| 2 | Supraorbital ridges | c.1 | 20 | Waist | a. | 164 | |
| 3 | Bony ear labyrinths | a. | 155 | 21 | Humerus | a. | 165 |
| 4 | Palate | a. | 156 | 22 | Femur | a. | 166 |
| 5 | Mandible | a. | 157,158 | 23 | Humero/femoral index | a. | 167 |
| 6 | Dental arcade | a. | 159 | 24 | Fingers | a. | 168 |
| 7 | Diastema | c. | 25 | Toes | a. | 169 | |
| 8 | Canines | c. | 26 | Iliac | a. | 170 | |
| 9 | Premolars | c. | 27 | Pelvis overall | d.3 | ||
| 10 | Incisors | c. | 28 | Knee | c. | ||
| 11 | Molars | b. | 29 | Valgus degree | c. | ||
| 12 | Saggital crest | b.2 | 30 | Talus | b. | 171 | |
| 13 | Canine tooth roots | a. | 160 | 31 | Foot | b. | 172 |
| 14 | Brain capacity—afarensis, graciles | a. | 32 | Meatus angle, face to cranium | a. | 173 (See Figure 15) | |
| 15 | Brain capacity—Robust types | a. | 33 | Overall size | b.4 | ||
| 16 | Brain capacity—Habilines | b. | 34 | Locomotion | b. | 174 | |
| 17 | Prognathicity | a. | 161 | 35 | Zygomatic arch | b.5 | |
| 18 | Shoulder | a. | 162 | 36 | Dental maturation | a. | 175 |
| Key: | The probable status conclusions about australopithecines and habilines for various characteristics have each been assigned into one of five groups (references are given where required): | |
| a = ape-like | ||
| b = doubtful, but leaning towards ape-like | ||
| c = indeterminate or irrelevant | ||
| d = doubtful, but leaning towards human-like | ||
| e = human-like | ||
| Notes: |
|
|
| Summary: |
|
|
| I have been generous towards the evolutionist position by allocating some features such as brow ridges and the knee to the indeterminate category. There still remain too many ape-type features, and too few human-type traits for us to accept that australopithecines (and habilines) are true intermediates. As can be seen, of the 36 features in question, only one can be allocated even to doubtful, leaning towards human-like, and only a few which can be put in the indeterminate group. The overwhelming impression is of a chimp-like creature, now extinct, or surviving as a variant of that creature. If australopithecines were truly bipedal, one would expect that there would be a much higher number of traits falling in the indeterminate (c), doubtful, leaning towards human-like (d), or human-like (e) groupings. | ||
Table 5. Summary of the morphological traits of alleged pre-human ancestors.
Summary and conclusions
The following facts seem reasonably well established:
- In maintaining that australopithecines are ape-like ancestors of human beings, the actual evidence leaves much to be desired. It is by no means a settled question—there is too much doubt and there are too many conflicting opinions regarding the significance of the various fragments. One would have expected these curious fossil creatures to have displayed many more human-type traits than they do (see Table 4).
- The degree of subjectivity involved has undoubtedly been a major factor—it is very obvious that the more prominent investigators have been too heavily influenced by the fame and fortune derived from the discovery of controversial fossils, and the subsequent published claims. In many cases the assertions made about the significance of the various fossils have been exaggerated in the first instance.
- New finds are always subject to later revisions, which often reverse or put in severe doubt the original claims made by the discoverers and their supporters in the heat of excitement surrounding the finds. The best approach is to maintain a healthy skepticism in regard to sensational early claims, and wait for the more independent researchers’ evaluations which are made in a more sober and dispassionate atmosphere. The latter are likely to be more accurate.
- The findings of the team led by Spoor appear to clinch the creationist argument that the australopithecines lie morphologically in the pongid line, and not in the human family line.
- We have the peculiar situation that there are now no evolutionary ancestors for the genus Homo, which includes H. erectus, H. neanderthalensis, and Homo sapiens. Neither are there any suitable evolutionary ancestors for the three modern African great apes, Gorilla gorilla, Pan troglodytes, and Pan paniscus, while on the other hand there are no modern descendants of the three very similar Pliocene and Pleistocene apes, A. boisei, A. robustus, and A. africanus. Table 5 indicates a more probable phylogeny for the higher primates, based on my studies.
- The media and the education establishment has played a dominant role in the indoctrination of the public, by focussing heavily on the initial and more sensational claims by palaeontologists, and by ignoring the more sober evaluations of later independent experts.
Acknowledgment
Carl Wieland is thanked for his invaluable help in drafting the text of this paper more clearly.
The late A. W. (Bill) Mehlert had a Diploma in Theology and lived in Brisbane, Australia. He was a keen student of Flood geology and the fossil record, including the supposed fossil evidence for human evolution, and wrote a number of important articles on these topics in the Creation Research Society Quarterly and Journal of Creation.
References
- Lewin, R., Bones of Contention, Penguin/Pelican, London, 348pp, 1989. Return to text.
- Reader, J., Missing Links, Penguin, London, p. 215 (citing Tobias, 1981), 1990. Return to text.
- Beasley, G.J., Pre-Flood giantism: a key to the interpretation of fossil hominoids and hominids, Journal of Creation 4:31, 1990 (citing Boaz and Tobias, 1983). Return to text.
- Lewin, ref. l, p. 129. Return to text.
- Susman, R. and Stern, J., The locomotor behavior of A. afarensis, American Journal of Physical Anthropology 60:279–317, 1983. Return to text.
- Shipman, P., Those ears were made for walking, New Scientist 143(1936):26–29, 1994. Return to text.
- Armstrong, S., South Africa—the missing pieces, New Scientist 143(1933):34, 1994. Return to text.
- Andrews, P., The descent of man, New Scientist 102:24, 1984. Return to text.
- Andrews, ref. 8, p. 24. Return to text.
- Bromage, T., Faces from the past, New Scientist January 11, p. 34, 1992. Return to text.
- Bromage, ref. 10, p. 35. Return to text.
- Johanson, D., Lucy, a 3.5 million-year-old woman shakes man’s family tree, Science ’81 2(2):50, 1981. Return to text.
- Gowlett, J., Ascent to Civilization, Alfred Knopf, New York, p. 27, 1984. Return to text.
- Tanner, N.M., On Becoming Human, Cambridge University Press, p. 180, 1981. Return to text.
- White, T.D., Suwa, S., Hart, W.K., Walter, R.C, WoldeGabriel, G., de Heinzellen, J., Clark, J. D., Aslaw, B. and Vrba, E., New discoveries of Australopithecus at Maka in Ethiopia, Nature 366:261–265, 1993. Return to text.
- Conroy, G.C. and Vannier, M.W., The nature of Taung dental maturation, continued, Nature 335:808, 1988. Return to text.
- Beynon, A. D. and Dean, M., Distinct dental development pattern in early fossil hominids, Nature 335:509, 1988. Return to text.
- Leakey, R. and Lewin, R., Origins Reconsidered, Little, Brown and Co., London, p. 155, 1992. Return to text.
- Leakey and Lewin, ref. 18, p. 156. Return to text.
- Leakey and Lewin, ref. 18, pp. 158–159. Return to text.
- Mann, A.E., Palaeodemographic Aspects of the South African Australopithecines, University of Pennsylvania Press, Philadelphia, p. 6, 1975 (Cited by Reader, Ref. 2, pp. 128–129). Return to text.
- Smith, H., On the dentition of the Taung skull, Nature 323:327–330, 1986. Return to text.
- Conroy, G.C. and Vannier, M.W., Dental development of the Taung skull from computerized tomography, Nature 329:625–627, 1987. Return to text.
- Mann, A.E., The nature of Taung dental maturation, Nature 323:123, 1988. Return to text.
- Wolpoff, M.H., Was Taung human or an ape? Nature 335:501, 1988. Return to text.
- Bromage, ref. 10, pp. 32–33. Return to text.
- Lewin, ref. 1, p. 78. Return to text.
- Gish, D.T., Evolution: the Challenge of the Fossil Record, Master Books, El Cajon, California, p. 178, 1985. Return to text.
- Johanson, D. and Edey, M. A., The Beginnings of Humankind, Penguin, London, p. 276, 1990. Return to text.
- Johanson and Edey, ref. 29, pp. 276–277. Return to text.
- Johanson and Edey, ref. 29, pp. 276–277. Return to text.
- Campbell, B., Humankind Emerging, 5th edition, Harper Collins, New York, p. 173, 1988. Return to text.
- Willis, D., The Hominid Gang, Viking Press, New York, p. 83, 1989. Return to text.
- Cherfas, J., Trees have made man upright, New Scientist 97:172–174, 1983. Return to text.
- Leakey and Lewin, ref. 18, pp. 194–195. Return to text.
- Cherfas, ref. 34, p. 172. Return to text.
- Johanson, D. C. and Shreeve, J., Lucy’s Child, Viking Press, London, 318 pp., 1990. Return to text.
- Leakey and Lewin, ref. 18, p. 193–194. Return to text.
- Zihlman, A., Pygmy chimps, people and the pundits, New Scientist 104:40, 1984. Return to text.
- Leakey and Lewin, ref. 18, p. 194. Return to text.
- Lovejoy, C.O., Evolution of human walking, Scientific American 259(5):86, 1988. Return to text.
- Cherfas, ref. 34, p. 172. Return to text.
- Leakey and Lewin, ref. 18, p. 118. Return to text.
- Leakey and Lewin, ref. 18, p. 118. Return to text.
- Reader, ref. 2, p. 188. Return to text.
- Johanson and Shreeve, ref. 37, p. 204. Return to text.
- Johanson and Shreeve, ref. 37, pp. 286–287. Return to text.
- Bunney, S., The fruits of walking on two legs, New Scientist 142(1925):18, 1994. Return to text.
- Johanson, D.C., Ethiopia yields ‘First Family’ of early man, National Geographic 150(6):790–810, 1976. Return to text.
- Cherfas, ref. 34, pp. 172–177. Return to text.
- Susman and Stern, ref. 5, pp. 279–317. Return to text.
- Leakey and Lewin, ref. 18, p. 195. Return to text.
- Willis, ref. 33, p. 245. Return to text.
- Johanson and Shreeve, ref. 37, pp. 194–201. Return to text.
- Johanson and Shreeve, ref. 37, p. 201. Return to text.
- Lowenstein, J. and Zihlman, A., False start of the human parade, Natural History 88(7):88, 1979. Return to text.
- Cherfas, ref. 34, p. 174. Return to text.
- Zihlman, ref. 39, p. 39. Return to text.
- Cherfas, ref. 34, p. 176. Return to text.
- McGowan, C, In the Beginning: a scientist shows why the creationists are wrong, Prometheus, Buffalo, p. 177, 1984. Return to text.
- Lovejoy. ref. 41, p. 86. Return to text.
- Cherfas, ref. 34, p. 173. Return to text.
- Susman and Stern, ref. 5, p. 291. Return to text.
- Cherfas, ref. 34, p. 174. Return to text.
- Gould, S.J., The upright ape, New Scientist 83(1171):739, 1979. Return to text.
- Lovejoy, ref. 41, p. 89. Return to text.
- Lovejoy. ref. 41, p. 83. Return to text.
- Campbell, ref. 32, p. 228. Return to text.
- Willis, T., Lucy goes to college, Bible-Science Newsletter 25(10):l–5, 1987. Return to text.
- Johanson and Edey, ref. 29, pp. 206–207. Return to text.
- Johanson and Edey, ref. 29, p. 207. Return to text.
- Susman and Stern, ref. 5, p. 298. Return to text.
- Cherfas, ref. 34, p. 173. Return to text.
- Cherfas, ref. 34, p. 173. Return to text.
- Cherfas, ref. 34, p. 173. Return to text.
- Bower, B., Hominids down to earth or up a tree? Science News 145(15):231, 1994. Return to text.
- Cherfas, ref. 34, p 172. Return to text.
- Shea, B.T., Knuckle walking ancestors, Science 259:293–294, 1993. Return to text.
- Cherfas, ref. 34, p. 176. Return to text.
- Cherfas, J., 1988. Personal communication. Return to text.
- Gish, ref. 28, pp. 167–168 (citing Oxnard, American Journal of Physical Anthropology 52:16, 1980). Return to text.
- Wood, B., Origin and evolution of the genus Homo, Nature 355:786, 1992. Return to text.
- Johanson and Edey, ref. 29, p. 249. Return to text.
- Johanson and Edey, ref. 29, pp. 254–255. Return to text.
- Johanson and Shreeve, ref. 37, pp. 196–197. Return to text.
- Johanson and Shreeve, ref. 37, p. 197. Return to text.
- Tuttle, R.H., The pitted pattern of Laetoli feet, Natural History 99(3):64, 1990. Return to text.
- Tuttle, R.H. and Webb, D., The patterns of little feet, American Journal of Physical Anthropology 78(2):316, 1989. Return to text.
- Tuttle, R.H. and Webb, D., Did Australopithecus afarensis make the Laetoli G footprint trail? American Journal of Physical Anthropology supplement S12:175, 1991. Return to text.
- Tuttle, ref. 87, p. 64. Return to text.
- Tuttle, R.H., Primate origins and evolution, American Journal of Physical Anthropology 85(2):244, 1991. Return to text.
- Oxnard, C.E., Lisowski, F.P. and Albrecht, G.H., Some African fossil footbones: a note on the interpolation of fossils into a matrix of extant species. American Journal of Physical Anthropology 41:191–216, 1974. Return to text.
- Oxnard, C, The Order of Man, Yale University Press, New Haven, p. 332, 1984 (cited by Gish,. D., Creation Scientists Answer Their Critics, Institute for Creation Research, El Cajon, California, p. 382, 1993). Return to text.
- Oxnard, ref. 93, pp. iii and iv. Return to text.
- Wood, ref. 82, p. 783 (Header). Return to text.
- Wood, ref. 82, p. 783 (citing Le Gros Clark, 1964 and Holloway, 1965). Return to text.
- Wood, ref. 82, p. 783 (citing J.T. Robinson, 1965 and C. L. Brace et al., 1972). Return to text.
- Wood, ref. 82, p. 784. Return to text.
- Wood, ref. 82, p. 785. Return to text.
- Wood, ref. 82, p. 785. Return to text.
- Wood, ref. 82, p. 787. Return to text.
- Lewin, ref. l, p. 145. Return to text.
- Lewin and Leakey, ref. 18, p. 161. Return to text.
- Wood, ref. 82, p. 788. Return to text.
- Johanson and Edey, ref. 29, p. 104. Return to text.
- Wood, ref. 82, p. 789. Return to text.
- Lewin, R., Human origins: the challenge of the Java skulls, New Scientist 142(1924):36–40, 1994. Return to text.
- Harris, M., Our Kind, Harper and Row, New York, p. 23, 1989. Return to text.
- Harris, ref. 108, p. 22. Return to text.
- Lewin, ref. 1, p. 160. Return to text.
- Gish, ref. 28, p. 166. Return to text.
- Beasley, ref. 3, p. 35 (citing R. Leakey, 1974). Return to text.
- Beasley, ref. 3, p. 35 (citing A. Walker, 1981). Return to text.
- Brace, C.L. and Montagu, A., Human Evolution, 2nd edition, MacMillan, New York, p. 271, 1977. Return to text.
- Brace, C.L. and Wolpoff, M., Allometry and early hominids, Science 189(4196):61–63, 1975. Return to text.
- Beasley, ref. 3, p. 36. Return to text.
- Lewin and Leakey, ref. 18, p. 119. Return to text.
- Beasley, ref. 3, pp. 32, 38. Return to text.
- Beasley, ref. 3, p. 43. Return to text.
- Lewin, ref. 107, pp. 36–40. Return to text.
- Holloway, M, Family matters, Scientific American 270(5): 15–16, 1994. Return to text.
- Groves, C., The origin of human species, Medical Journal of Australia 151:681, 1989. Return to text.
- Reader, ref. 2, p. 189. Return to text.
- Shipman, ref. 6, pp. 26–29. Return to text.
- Shipman, ref. 6, p. 29. Return to text.
- Bower, ref. 76, p. 231. Return to text.
- Mehlert, A.W, A review of the present status of some alleged early hominids. Journal of Creation 6(1):10–41, 1992. Return to text.
- Shipman, ref. 6, p. 29. Return to text.
- Johanson and Shreeve, ref. 37, p. 201. Return to text.
- Cherfas, ref. 34, p. 174. Return to text.
- Brace and Montagu, ref. 114, p. 271. Return to text.
- Clark, G., World Prehistory in Perspective, 3rd edition, Cambridge University Press, pp. 5, 22, 1977. Return to text.
- Bower, B., Fossils put new face on Lucy’s species, Science News 145:212, 1994. Return to text.
- Lewin, ref. 1, p. 48. Return to text.
- Eldredge, N., The Miner’s Canary: Unravelling the Mysteries of Extinction, Prentice-Hall, New York, p. 184, 1993. Return to text.
- Johanson and Edey, ref. 29, p. 24. Return to text.
- Bunney, S., Will the real Homo habilis stand up? New Scientist 120(1636):33, 1986. Return to text.
- Hummer, C., A plea for caution about skull 1470, Creation Research Society Quarterly 14:168–172, 1977. Return to text.
- Johanson and Edey, ref. 29, p. 146. Return to text.
- Bromage, ref. 10, pp. 32–35. Return to text.
- Bromage, ref. 10, p. 33. Return to text.
- Bromage, ref. 10, pp. 32–34. Return to text.
- Lubenow, M., From Fish to Gish Creation Life Publishers, El Cajon, California, p. 208, 1983. Return to text.
- Cherfas, J. and Gribbin, J., Descent of man or ascent of ape? New Scientist 91:592–595, 1981. Return to text.
- Beasley, ref. 3, pp. 5–55. Return to text.
- Oxnard, ref. 93, p. 320. Return to text.
- Begley, S., Out of Africa: a missing link, Bulletin/Newsweek 116(5940):76–77, 1994. Return to text.
- Begley, ref. 147, p. 76. Return to text.
- Cowley, G. and Salzhauer, A., Humankind’s first steps, Bulletin/Newsweek 116(5985):65, 1995 (diagram). Return to text.
- Cowley and Salzhauer, ref. 149, p. 65. Return to text.
- Anonymous, The age of australopithecines, Nature 372:31, 1994. Return to text.
- Ridley, M., Who doubts evolution? New Scientist 90(1259):830–832, 1981. Return to text.
- Johansen and Edey, ref. 29, pp. 276, 277. Return to text.
- Campbell, ref. 32, p. 173. Return to text.
- Shipman, ref. 6, pp. 36–39. Return to text.
- Johanson and Edey, ref. 29, p. 274. Return to text.
- Johnson and Shreeve, ref. 37, p. 86. Return to text.
- Johanson and Edey, ref. 29, p. 273. Return to text.
- Johanson and Edey, ref. 29, p. 271. Return to text.
- Johanson and Edey, ref. 29, p. 275. Return to text.
- Bromage, ref. 10, p. 33. Return to text.
- Cherfas, ref. 34, pp. 172–173. Return to text.
- Leakey and Lewin, ref. 18, pp. 193–196. Return to text.
- Leakey and Lewin, ref. 18, p. 194. Return to text.
- Leakey and Lewin, ref. 18, p. 118. Return to text.
- Leakey and Lewin, ref. 18, p. 118. Return to text.
- Leakey and Lewin, ref. 18, pp. 118–119. Return to text.
- Cherfas, ref. 34, p. 172. Return to text.
- Cherfas, ref. 34, pp. 174, 177. Return to text.
- Cherfas, ref. 34, p. 173. Return to text.
- Cherfas, ref. 34, p. 176. Return to text.
- Cherfas, ref. 34, pp. 174–176. Return to text.
- Bromage, ref. 10, p. 33. Return to text.
- Cherfas, ref. 34, pp. 172–178. Return to text.
- Bromage, ref. 10, p. 33. Return to text.



Readers’ comments
Comments are automatically closed 14 days after publication.