

Journal of Creation 26(3):60–67, December 2012
Browse our latest digital issue Subscribe
Vas deferens—refuting ‘bad design’ arguments
The vas deferens is an important part of the male reproductive system. However, some anti-creationists have recently criticized its route for being too indirect, thus something which no engineer would design. However, anatomists have already given good reasons for this structure, including the increased flexibility of the testes to move toward and way from the body to regulate temperature. Critics have also overlooked engineering considerations, providing enough length to build up power and to mix the essential ingredients of semen, and to avoid ‘ovalling’ (kinking in a soft pipe when bending).

The vas deferens (Latin for ‘carrying-away vessel’; plural: vasa deferentia), also often referred to, especially in medical literature, as the ductus deferens, is a vital part of the anatomy for many male vertebrates. It stores and transports the sperm from the testis to the penis for insemination, in reproduction. Without it, the species would be extinct (some men voluntarily have themselves sterilized by having this tube cut and having the cut ends sealed—a vasectomy—or nowadays having these ducts blocked to prevent sperm from passing).
Yet, like a few other features, evolutionists have attacked its looping indirect route as ‘bad design’. A good example is Richard Dawkins:
“The vas deferens is the pipe that carries sperm from the testes to the penis … . It takes a ridiculous detour around the ureter, the pipe that carries urine from the kidney to the bladder. If this were designed, nobody could seriously deny that the designer had made a bad error.”1
Dawkins concludes in an almost arrogant manner that:
“Examples like this must surely undermine the position of those who hanker after ‘intelligent design’.”2
A bad claim about bad design
A fundamental consequence of something having been badly designed is that it functions poorly because of this particular design. However, Dawkins and his type of evolutionist merely assert that no engineer would create a vas deferens with such a detour, and that this in itself is evidence for bad design. They fail to demonstrate how this detour of the vas actually harms or disadvantages the male reproductive system. This system actually works quite well; and if it performed poorly because of this, then, by the evolutionists’ own reasoning, natural selection would have punished this ‘design’. Rather, this is a fallacious argument from personal incredulity, something Dawkins affects to despise, despite his own use of it.
We should also learn from previous faulty claims of ‘bad design’ by Dawkins, including the backwardly wired retina, prostate, and recurrent laryngeal nerve. Analysis beyond Dawkins’ superficial assessment proves that these organs are very well designed, and that Dawkins’ alternative proposals would be much worse.3 As will be shown, this is true of the vas as well.
Some background
Embryology
Many features we observe are not the result of evolutionary history, but embryological development. This is also so with the vas deferens. After all, multi-cellular animals start off as a fertilized egg (the zygote), which then becomes an embryo. For the purposes of producing an adult organism, the needs of the growing embryo are just as important as those of the adult.
Human embryos begin as morphologically sexually dimorphic (i.e. containing characteristics of both sexes). This is so, because they all have basically the same genetic information, and this information is expressed as efficiently as possible as the embryo develops. This is design economy. For example, in all human embryos, at first both the Müllerian duct system (female) and the Wolffian duct system (male) develop, because both sexes have the genetic information for these structures. The differentiation which occurs later is the result of designed chemical signals that control the expression of the information. The male sex-determining gene is found on the Y chromosome4 which controls the levels of testosterone and dihydroxytestosterone (DHT) secretions. Above a certain level, these hormones suppress the development of the Müllerian duct system, which then later degenerates in males, and promote the Wolffian duct system, so masculine characteristics develop. Indeed, the vas deferens is one of the structures that forms from the Wolffian duct or mesonephric duct.5 Below a certain hormone level, the opposite happens, and the embryo starts to develop the female structures.6
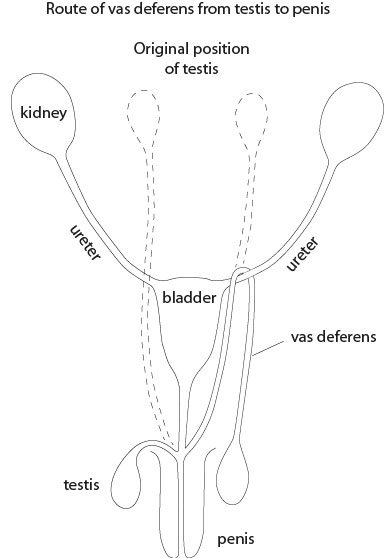
This dimorphism is also responsible for the testes and ovaries developing from the same structure, the genital ridge, close to the starting point of the kidneys. In males, the testis is joined to the scrotum via a cord called the gubernaculum testis. This cord doesn’t grow, so as the baby grows and the scrotum moves further away, it drags the testis with it, eventually through the body wall7 (for reasons explained below). The vas deferens just follows this route.
Ontogeny and phylogeny
It should be clear from the above information that the course of the vas deferens is a result of the embryonic development(ontogeny), not the evolutionary development or phylogeny (contra Dawkins), just like male nipples, for example.
The invention of the idea that evolutionary history parallels embryonic development, or the ‘biogenetic law’, was made by the fraudulent Ernst Haeckel (1834–1919). Even after repeated refutations, there are still those trying to rehabilitate Haeckel to shore up their evolutionary belief system.8,9 Sadly, the faulty idea of ‘ontogeny recapitulates phylogeny’ still has an unhealthy hold on people’s minds.

In embryonic development, the kidneys and both the urinary and genital ducts are closely interconnected and related. In amniotes in general,10 and humans in particular, three nephric (kidney) structures succeed one another. They are the pronephros, mesonephros (hence modern reference to the mesonephric duct), and the metanephros. These structures come with their associated duct systems, which differ from one another. Not only embryonic fishes and amphibians (and briefly reptile, bird and mammal embryos) but also adult hagfish and some bony fish species display the pronephros setup. So unsurprisingly, some sources state or at least hint strongly11 that this is to some extent related to the evolution of the nephric system. However, a widely distributed textbook on physiology of the vertebrate body says:
“It is often stated or implied that these three are distinct kidneys that have succeeded one another phylogenetically [in the creature’s evolutionary history] as they do embryologically. However, there is little reason to believe this. The differences are readily explainable on functional grounds; the three appear to be regionally specialized parts of the holonephros, which serve different functions.”12
The authors go on to explain:
“An actively growing embryo has wastes to excrete. When kidney tubules are formed anteriorly, they begin the process of urine formation. The formation of a urinary tube for drainage cannot be delayed until the entire kidney is formed; the anterior tubules just cannot wait that long.”12
So, a simpler kidney-and-duct system is needed long before it would actually be practically possible to have a fully formed metanephric system. So the simpler system has nothing to do with evolutionary history, and everything to do with the requirements of the embryo at the early stage. This is practical design economy once again.
Zoological purpose
There is only a narrow temperature range in which healthy male human gametogenesis (sperm formation) can occur. One important zoological reason for the vas deferens taking the course it takes, is the fact that the sperm is so temperature dependent. In order to regulate temperature, the testes move closer or further away from the body (in the scrotum itself). They will move closer in order to raise temperature, but away to lower temperature. The scrotum and testes are able to do this with a system involving the cremaster muscle. The cremaster surrounds the testis, and contracts when exposed to cold (or during sexual arousal to protect the testis), to bring it closer to the body for warmth. Warmth causes it to relax so that the testis can move further away.13,14 (The thermoregulation system also involves the tunica dartos, which lies beneath the skin of the scrotum.)
With a vas taking the much shorter routes (as figure 1 attempts to illustrate), or a direct route out to the penis, as Dawkins alleges “any sensible engineer would design”, this important cremaster function would not be possible. But with its existing arrangement, there is enough tolerance and the testes can literally be ‘given some rope’, to move closer to or away from the body.
The above reason was outlined to me by my own vertebrate zoology professor. To my surprise, when I asked him about this supposedly odd arrangement, he did not go on with the whole evolutionary story, but explained this practical purpose, even though he is an evolutionist.
This is one of many examples of how evolution is useless for real biology. Rather, this evolutionist asked the same questions as a creationist would: what functions could this structure enable? Also, it shows that Dawkins is out-of-step even with evolutionary experts in the fields he is talking about.15
Engineering and more biological considerations
There are more possible explanations for the course of the vas deferens from an engineering, practical and biological point of view.
Why can’t the vas deferens lead directly to the penis?
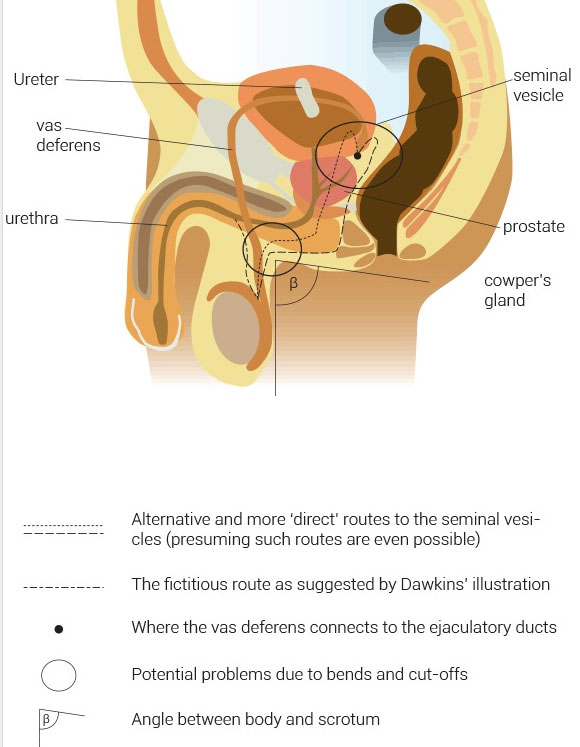
Lay people might ask themselves this seemingly obvious question. But (surprisingly, given that he is eminently qualified in biology), Dawkins’ illustration (see figures 1 and 3) naïvely suggests this as the most direct route to the penis (and implies that this is a better option for a more direct route). He hints in the text that any ‘sensible engineer’ would ‘reroute’ the vas deferens like this. So why must the vas lead to the ejaculatory ducts close to the bladder? One reason is that semen is actually composed of several ‘ingredients’, some of which must be added by various glands, which sit along the way of the ejaculatory duct and urethra. These include the seminal vesicles, the prostate gland and Cowper’s gland/the bulbourethral gland. Only after all these have made their contribution is the final product of semen ready for its purpose.
More problems with more direct routes
Volume and infertility
An interesting factor comes to mind when we think of infertility in males. Normal male semen generally has more than 100 million sperms per millilitre (ml). Much variation can occur among individuals, but the rule is more or less that:
- individuals with at least 20 million sperm/ml, or a total of 50 million per ejaculation, are likely to be fertile
- those with 10 million spermatozoa/ml or less are likely to be sterile.16
How does this fit in with the benefits of a longer vas, or the disadvantages of having a shorter straight-routed vas, which Dawkins’ diagram suggests? The answer is in volume. A shorter vas deferens would have a smaller volume. A shorter vas with smaller volume would also mean that (far) less sperm per volume unit ejaculation could be delivered. This means that with Dawkins’ significantly shorter proposed vas, the sterility rate of males would likely be much higher. And under his evolutionary scenario, they would not leave as many offspring, so this trait would be unlikely to persist.
More energy, more work done, higher ‘power’
For any fluid to travel between point A and point B, there needs to be a difference in pressure or Δp between point A and B. If we want our fluid to travel from A to B, we need to have a higher pressure at A than B. This is sometimes referred to as a ‘pressure gradient’ in literature. This refers to the change in pressure over a distance, or:

In thermal fluid networks, however, we would use a pump with a specific ‘power’ to create a pressure differencein order for the fluid to be transported. Different pumps with different performance curves and power would be used for different applications. In a network with higher resistance and losses, or alternatively, where we need a higher exit velocity (or larger volumes of water) compared to a network which has lower resistance, losses and the other properties, we would carefully choose a pump with higher ‘power’ (and a more suitable performance curve), where the power is defined as the work done by the pump per unit time, or:

If there were a human pump at the end of a static, and a hard, rigid pipe like PVC, Dawkins’ claims might make sense. Instead the vas is a flexible duct with relatively thick surrounding muscles. These are responsible for creating the necessary Δp or pressure rise in the vas deferens and adding the energy to the fluid, via peristaltic waves (see below). The important question then becomes: how would we define the work done (and ultimately the power) by the peristaltic wave in the vas deferens on the semen? The meaningful way to express this seems to be the old and most familiar definition:
“The work done on a particle is equal to the force acting on the particle in the direction of movement (in this case, the streamline), multiplied by the distance the particle travels.”17
In mathematical terms we can write it as follows.
For a variable force, we write:

where F is the net force or component thereof (Fcosθ with θ the angle between the direction of travel and the force) acting on the body or particle in the direction of the travel, and r1 to r2, or s1 to s1, the finite path the particle travels.
Alternatively, to simplify a bit, for a constant force acting on a body or particle parallel to its direction of motion, we would write:

where d is the distance the body or particle travels (and F, the same as before).
Having defined the above, we can now perhaps try to define the ‘power’ of the vas deferens. Knowing that the power is defined by the work done divided by a chosen time interval, we can now substitute the formula(e) for work into the power formula. Thus:

Again, to simplify a bit, we can write for a constant force:

Having defined the above, we can now look at why the longer vas deferens may exhibit interesting functions. If we now compare a short vas and a longer vas, we can see that, for the same numerical value of Δt, or the same time interval, and the same numerical value of the force F, which the peristalsis can possibly produce, we can actually get a larger numerical value for the d-term, because the vas is longer. This means that we will obtain a higher power generation with a longer vas, as illustrated by figure 2. On the other hand, if we have a vas deferens as Dawkins frivolously suggests in his illustration (see figure 3), which is, let’s say for example, four times as short as the actual vas, we will need a force F four times the numerical value, in order to obtain the same power generation. For the same vas with the same muscle, just shorter, this might just not be possible. So the longer vas deferens means that we have a higher power pump, and the seminal fluid has more kinetic energy. This section shows that Dawkins has erred by treating a dynamic feature as a static one, a mistake he has made before (with the eye).18
Why we would need higher power and more kinetic energy?
Since the energy/work/power of the vas deferens is a new consideration, the precise or practical purpose of it may be yet to be discovered. However, there are a few possible theoretical advantages which we can discuss, which seem to some extent obvious.
Better mixing of the semen
As we have already discussed, the final semen/ejaculate is composed of several products from several glands. The seminal vesicles contribute about 60% of the total volume of the semen.19 When the spermatozoa coming from the vas deferens and the ejaculate coming from the seminal vesicles (as well as the other sex glands) meet and mix, there may be an advantage to spermatozoa having a higher velocity and more kinetic energy. In short, it can possibly ensure that the spermatozoa mix better with the products of the other sex glands, but especially that of the seminal vesicles, so that the semen ends up as more homogenous. There are good reasons why this is necessary, as a medical expert has pointed out:20
- The distal part of the epididymis and the vas contain all the spermatozoa in only 10% of the ejaculate. Therefore good mixing is essential for all the spermatozoa to become active.
- The seminal vesicles contract just after the vasa (which contract in concert with the prostate), ensuring as far as possible that the urethra is emptied of all sperm. The seminal vesicular secretions contribute about 60% of the ejaculate, the prostate about 30%, with small contributions from other glands entering directly into the prostatic urethra, all in the same area. Any other proposal for re-routing the vas must allow for effective mixing. The constituents of the semen are highly complex and are required to perform multiple sequential functions in order to deliver actively viable spermatozoa to the right place at the right time.
- The sperm are inactive on account of multiple factors present in the fluids of the vas, mainly its pH (moderately acidic). They become active in a neutral or mildly alkaline environment provided by mixing with the alkaline fluids of the prostate and seminal vesicles. The pH of the ejaculate is about 7.5, slightly alkaline. So on this point, proper mixing is essential.
- The vaginal secretions are normally acidic (pH 3.5–4.0). So unless the sperm are well protected by mixing with the seminal fluids, they will suffer excessive attrition before being deposited in the seminal coagulum, near the cervix.
Ejaculation
The overall process of ejaculation is relatively complex, and consists of several sub-mechanisms, each acting at the right place and time. The process can be divided into several steps. For example:
“Emission: This is a sympathetic response where semen is deposited to the prostatic urethra via the ejaculatory ducts, just after peristalsis takes place in the vas deferens.
“Ejaculation: This is the phase where the semen is expelled through the external urethral orifice (after having travelled through the urethra). This process involves several mechanisms, such as the periurethral muscles (muscles encircling the urethra) performing a series of rhythmic, involuntary contractions, which expel the semen. The vesicle sphincter at the neck of the bladder also closes tightly during this process in order to prevent a ‘backwash’ of the ejaculate.”21,22
From the above one can see that the overall process of ejaculation is not achieved by one single mechanism, but a group of functions working together.
However, there is a possibility that the higher power of the vas, giving the spermatozoa a higher exit velocity, can assist the semen being properly deposited into the urethra during the phase of emission.
Potential cut offs in more viable direct routes
What then, about routes which do connect at the ejaculatory ducts, but are still more direct in their paths, and do not make such loops? (See fictitious suggestions in figure 1) Even in this approach, we can theoretically point out some flaws and noticeable problems. Of course, this also dismisses the deliberate benefits which the long vas may have, as pointed out above.
Some background
In thermal fluid networks that engineers work with, such as those that would typically occur at mines and plants, pipes are mostly used which are made from hard and rigid materials such as PVC, steel, concrete and so forth. However, the ‘tubes’ and ducts in our body, including the vas deferens, are soft, flexible tubes. This must also be the case so that the contents of these can be transported and manoeuvred via peristaltic waves (see below). This all works well, but there is another factor which needs to be considered.
If we take something like a domestic hose pipe, commonly used in our gardens, and bend it from its straight line of 180° to say a right angle (90°), what will happen? Will water still be able to flow through our hose pipe? Can you drink out of a highly bent straw?
The answer is, no, hardly. This is because the hose, like ducts in animals, is made of a softer material, and as these pipes are bent, the cross-sectional area at the bend decreases, which in turn increases the resistance and ultimately restricts the liquid from flowing. Soft pipes have this problem of decreasing cross-sectional area with bending, which can lead to cut offs. This is known in engineering circles as ‘ovalling’.
In hard-material pipes, preproduced bends such as ‘elbows’ are inserted, so the cross-sectional area remains constant, and thus do not cut off the water (though interestingly enough, these also often increase so-called secondary losses).
The vas deferens would have exactly the same ‘ovalling’ problem as other artificial soft tubes. It is different with very bendy soft tubes in our bodies like the small intestines, because they have structures analogous to the premanufactured man-made ‘elbows’.23 But this would not work well with the vas, as the vas gets pulled a bit back and forth with the moving of the testes. Thus there would be a trade-off with the flexibility required for thermoregulation. Furthermore, the vas has gotten the highest muscle-to-lumen ratio of any hollow viscus (plural: viscera) in the body. This means that the thick muscle body wall (see figure 4) of the vas would very easily and quickly close or cut off the lumen, through which the fluid must move, in the vas deferens, when the vas bends significantly.
Two hypothetical more direct routes clearly show problems in regards to cut-offs, with at least two hypothetical cut-off places, where the vas can potentially close due to bending. (See figure 1). During intercourse, it is a possibility that the angle β (in figure 1) will decrease, creating a bent tube with the cut-off problem.
But the actual vas in its existing position is remarkably straight, and avoids this problem altogether! Even a smaller β is not a problem at all with the existing vas design.
Obstruction of the peristaltic wave
A peristalsis is a muscular action which moves semisolid matter in the human body from one place to another; for example, through the alimentary canal. This occurs via a peristaltic wave which contracts (and closes) behind the matter, and then relaxes on the other side while moving along the duct. This then forces the matter along the duct. Peristalsis is also responsible for transporting the semen in the vas deferens to the ejaculatory ducts.
We have already seen that other, more direct, routes taken by the vas deferens would cause bending. I suggest that such bends, even if not entirely closed, may indeed either obstruct or significantly weaken the peristaltic wave. This is because it may interfere with the ability to close and open via contraction and relaxation at the right place, in the right sequence and at the right time.
Testing the testes hypothesis of evolutionists
Another evolutionist who has tried to make the anatomy of the testes and vas deferens fit an evolutionary framework, is none other than Steve Jones, the British anticreationist geneticist. In December 2011, he wrote an article for the Daily Telegraph (UK) complaining about many of his students not accepting Darwinian evolution. He wrote:
“In those happy days [when our ancestors were fish] the testes were deep within the main body mass, close to the liver (as they still are in our marine cousins). They were connected to the outside world by a pair of straight tubes. Then came the move on to land and the shift from cold blood to warm. That had lots of advantages, but faced the unfortunate male with a problem, for the delicate machinery for making sperm works best at low temperatures, perhaps to reduce the number of errors made as DNA is copied.
“The solution was a messy compromise in which the testes migrated south and emerged in their present form in an elegant external sac (which makes them, as I never fail to point out to students, both literally and figuratively the coolest part of any man’s body).”24
There are several problems with these hand waving proposals. Firstly, one wonders how the testes would know in advance over a period of time that they had to ‘migrate south’ in order to eventually be capable of exiting the body in order to contribute to fertilization. After all, evolution is a blind and purposeless process that has no eventual goal, and testes being “deep within the main body mass, close to the liver” would have had to have a goal to eventually ‘get out’ into a scrotal sac. So Jones himself displays ignorance of Darwinian evolution’s non-goal-directedness/purposelessness.
But there is another proverbial spanner in the wheel. Certainly the monotremes, allegedly primitive egg-laying mammals, have intra-abdominal testes (thus not in a scrotal sac outside the body at all). However, so do a number of placental mammals—both small ones such as shrews, hedgehogs, and moles, and some very large mammals, such as the hippopotamus and elephants.25 How, then, if all mammals evolved from a common ancestor, can temperature be the driving selective force (as if that alone is sufficient anyway) for the testes to descend outside of the body, if there are clear examples of mammals doing just fine with warm internal testes?
Of course, creationists do not hold to the ideas of either common ancestry for all mammals or that these structures developed by purely natural processes de novo. Thus we know that spermatogenesis in most mammals is indeed inhibited by higher temperatures. However, from a creationist point of view, that a handful of mammals have internal testes presents no paradox: all mammals did not evolve from a common ancestor. One possibility is that God created a select few mammals with internal testes, where higher temperature is not an issue, in order to serve the purpose of damaging a neat evolutionary story.26 This problem may not be lost on Dawkins, as he is careful to write (emphasis added):
“… [when] the testes descended to their current position in the scrotum (for reasons that are unclear, but are often thought to be associated with temperature) … .”27
If Dawkins is uncertain of the reason the testes descended to their current position then he cannot be quick to attack non-evolutionists on this point, or assert that ‘no designer would have designed it this way’. He does not even seem to know why ‘evolution’ supposedly did it.
And wrong demonstration too …
As if Dawkins and Jones do not already display enough ignorance, there is one last critique to be made of Dawkins. In his book, the Greatest Show on Earth, where he makes his claims, Dawkins gives his readers an illustration which is supposed to show how “evolution simply kept on lengthening the vas at only a very small marginal cost” (see figure 3). The illustration looks neat, but is actually incorrect, which undermines the neatness of the evolutionary story the picture tries to ‘paint’ in the readers’ mind. In Dawkins’ illustration the vas deferens connects up in the penile urethra (the part of the urethra going through the penis). This is not where the vas deferens unites with the urethra at all. Instead, it connects up high behind the bladder at the seminal vesicles and above the prostate. The vas actually never connects up to the urethra per se directly, (let alone the penile urethra). Of course, this is not the most serious charge against Dawkins, but had any creationist made such an error in an argument they tried to make, evolutionists would have likely waved it around as evidence for creationists’ ignorance of anatomy. So it is quite fair to point it out.
Conclusion
It should be clear that the charges made against the design of the vas deferens are quite frivolous, and so is the suggested alternative by Dawkins. Not only is the vas deferens a legacy of necessary embryonic development, but there are even more interesting advantages to its design. Some of these are straightforward, and reasonably well known to anatomists. Other proposals raised here involving fluid mechanics are new. But even if some may require revision at a later time, others have been long known to informed evolutionists and are strong enough that the overall picture would be unaffected. It is hoped that the combined arguments will stimulate much thought by fellow creationist scientists in order to further refute such ‘bad claims for bad design’ for other structures, but also in regards to the vas deferens. Claims that the vas is badly designed, or even just a legacy of evolutionary history, do not hold up under close investigation and scrutiny.
Acknowledgements
The author would like to thank the following persons: Dr Jonathan Sarfati for his significant contribution to this paper and persistent encouragement to write and complete this. Without his contributions, this paper would likely not have been possible. Thanks also to Dr Carl Wieland for his important medical insights and checks as well as Dr Andrew Hodge.
References
- Dawkins, R., The Greatest Show on Earth: The Evidence for Evolution, Free Press, New York, p. 364 2009. Return to text.
- Dawkins, ref. 1, p. 365. Return to text.
- See for example Sarfati, J., The Greatest Hoax on Earth? Refuting Dawkins on Evolution, ch. 15, Creation Book Publishers, Atlanta, GA, 2010. Return to text.
- Mitchell, B. and Sharma, R., Embryology: An illustrated Colour Text, Elsevier/Churchill Livingstone, Philadelphia, PA, p. 51, 2005. Return to text.
- Langman, J., Medical Embryology, 3rd edn, Williams and Wilkins, Baltimore, MD, p. 181, 1975. Return to text.
- Bergman, J., Is the human male nipple vestigial? J. Creation 15(2):38–41, 2001. Return to text.
- Langman, ref. 5, pp. 188–189. Return to text.
- Such as Richards, R.J., Haeckel’s Embryos: Fraud not Proven, Biology and Philosophy 24:147–154, 2009 | DOI 10.1007/s10539-008-9140-z; Published online: November 2008. Return to text.
- van Niekerk, E., Countering revisionism—part 1: Ernst Haeckel, fraud is proven, J. Creation 25(3):89–95, 2011. Return to text.
- Amniotes are a group of vertebrates where the embryo and fetus are suspended in a fluid-filled sac called an amnion. Amniotes include reptiles, birds, and mammals. Return to text.
- For example: Hickman, C.P., Roberts, L.S., Keen, S.L., Larson, A., I’Anson, H. and Eisenhour, D.J., Integrated Principles of Zoology, 14th edn, McGraw–Hill, Boston, MA, p. 673, 2008. Return to text.
- Romer, A.S. and Parsons, T.S., The Vertebrate Body, 6th edn, Saunders College Publishing, Philadelphia, PA, p. 407, 1986. Return to text.
- Jindal, S.K., Introduction to Animal Physiology, p. 198, New India Publishing Agency, New Delhi, 2011. Return to text.
- Van Der Graaf, K.M. and Fox, S.I., Concepts of Human Anatomy and Physiology, 4th edn, WCM /McGraw-Hill, Boston, MA, p. 845, 1999. Return to text.
- For another example, see Venter vs Dawkins on the Tree of Life—and Another Dawkins Whopper, evolutionnews.org, 9 March 2011. Dawkins was making his common false claim that the genetic code is completely universal. Craig Venter, of artificial life fame (see creation.com/venter), points out “the Mycoplasmas (the tiniest self-reproducing bacteria) use a different genetic code that would not work in your cells”. Dawkins was totally taken by surprise. Return to text.
- Moore, K.L. and Persuad, T.V.N., The Developing Human—Clinically Orientated Embryology, 8th edn, Saunders Elsevier, Philadelphia, PA, pp. 29–31, 2008. Return to text.
- Munson, B.R., Young, D.F., Okiishi, T.H. and Heubsch, W.W., Fundamentals of Fluid Mechanics, 6th edn, SI Version, Wiley, New York, p. 103, 2010. Return to text.
- See Sarfati, ref. 3, p. 269. Return to text.
- Guyton, A.C. and Hall, J.E., Textbook of Medical Physiology, 10th edn, Saunders Elsevier, Philadelphia, PA, pp. 916–922, 2000. Return to text.
- Thanks to Dr Andrew Hodge for providing this information. Return to text.
- Moore, K.L. and Persuad, T.V.N., The Developing Human—Clinically Orientated Embryology, 8th edn, Saunders Elsevier, Philadelphia, PA, p. 28, 2008. Return to text.
- Roberts, M. and Jarvi, K., Steps in the investigation and management of low semen volume in the infertile man, Can. Urol. Assoc. J. 3(6):479–485, December 2009; ncbi.nlm.nih.gov/pmc/articles/PMC2792416/; accessed 30 May 2012 Return to text.
- Also, with the small intestine, the matter moving through it is of a different nature/constituency, it moves much more slowly, and also uses another gut movement in addition to peristalsis, called segmentation. The latter is distinct from peristalsis in that it constantly divides and squeezes the food, moving it back and forward. See Hickman et al. ref. 11, p. 714. Return to text.
- Jones, S., Islam, Charles Darwin and the denial of science. A growing number of biology and medical students are rejecting the very basis of their chosen subject in favour of creationism, The Daily Telegraph, 3 December 2011; telegraph.co.uk/, accessed 7 March 2012. Return to text.
- Sodera, V., One Small Speck to Man—The Evolution Myth, 2nd edn, Vija Sodera Productions, p. 110, 2009. Return to text.
- This explanation is on the lines of Walter ReMine’s biotic message theory. That is, the evidence from nature points to a single designer, but with a pattern which thwarts evolutionary explanations. See ReMine, W.J., The Biotic Message: Evolution Versus Message Theory, Saint Paul Science, Saint Paul, MN, 1993; see review: Batten, D., J. Creation 11(3):292–298, 1997; creation.com/biotic. Other creation scientists are welcome to come up with other suggestions and interpretations. Return to text.
- Dawkins, ref. 1, pp. 364–365. Return to text.
- Kerr, J.B., Atlas of Functional Histology, Mosby, London, p. 357, 1999. Return to text.

Readers’ comments
Comments are automatically closed 14 days after publication.