

Journal of Creation 16(3):84–88, December 2002
Browse our latest digital issue Subscribe
An objective ancestry test for fossil bones
By the look of the curve of a fossil toe bone and the slant of its joint surface, recent reports concluded that it was from an ancestor of apes and humans, Ardipithecus ramidus kadabba, that walked on two legs. The question arose as to whether there might be a simple method yielding objective evidence to bridge the gap between those scant subjective determinations and that far-reaching conclusion. Accordingly, an objective, valid, reliable and calibrated correlational method of substantiating that conclusion was devised and successfully tested. For monkey (baboon), ape (chimpanzee) and human, similar were the ape and monkey, dissimilar were the human and monkey and most dissimilar were the ape and human. The monkey and ape similarities to human bone were less than for an anatomically different bone. The fossil toe bone had scant similarity to humans, dissimilarity to monkeys and most dissimilarity to apes with the dissimilarities to monkeys and apes like those for an anatomically different bone. The results of this objective ancestry test contradicted the conclusion that Ardipithecus ramidus kadabba was an ancestor of apes and humans that walked on two legs. Instead, these objective analyses provided evidence that apes are similar to monkeys, but monkeys and apes have no similarity to humans.
Amongst other fossils, Haile-Selassie reported a left, fourth digit, phalanx 1, toe bone 31.9 mm long designated Ardipithecus ramidus kadabba, AME-VP-1/71, 5.2 Ma. He reported:
‘AME-VP-1/71 shows a mosaic of features shared with both apes and A. afarensis. The proximal pedal phalanges of A. afarensis are unique in combining both strong phalangeal curvature (similar to apes) with a dorsally canted proximal joint surface (similar to later hominids). The dorsal orientation of this surface in AME-VP-1/71 may therefore constitute important evidence of a unique pedal morphology in this specimen similar to that in Hadar.’1
The interpretation given by Robinson was, ‘This toe bone (AME-VP-1/71) proves the creature walked on two legs. … How apes became human. Ardipithecus ramidus kadabba. What a new discovery tells scientists about how our oldest ancestors stood on two legs and made an evolutionary leap.’ His artist’s conception on the cover of Time was an upright chimpanzee-like creature with blue eyes.2 Except for the blue eyes, it bore a striking resemblance to the Piltdown forgery of 1912–1952.3
It is not clear that length was the only objective measurement made on AME-VP-1/71, but if it was, then the question arose as to whether there might be a simple method to yield objective evidence to bridge the gap between those scant subjective determinations and that far-reaching conclusion.
Accordingly, an atlas of primate gross anatomy by Swindler and Wood was obtained to scan the renderings of the comparable bone in baboons, chimpanzees and humans.4 If AME-VP-1/71 tested intermediate between chimpanzees and humans and resembled least baboons, then the conclusions of Haile-Selassie and Robinson would have objective support. The conceptual design of this test was to objectively determine for each bone a central axis and the deviations from it to the lateral and medial bone borders at decile distances from the proximal to the distal end. For each bone in the same order, these deviations in tenths of millimetres would be tabulated and correlated. A high correlation between two bones would indicate a high degree of similarity.
Methods
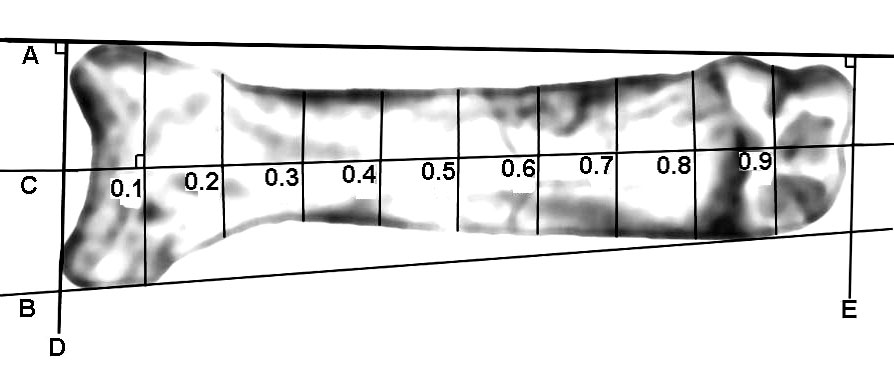
For a plantar or dorsal view, from proximal to distal, a straight sideline was drawn joining the lateral-proximal extreme of the bone with the lateral-distal extreme. The same was done for the medial extremes. See A and B of Figure 1.
To determine the central axis, a straight line was drawn perpendicular to the medial sideline such that it provided a tangent to the proximal extreme of the bone then crossed the lateral sideline. See D of Figure 1. The same was done for the distal extreme of the bone. See E of Figure 1. Between A and B, D and E were divided in half to determine the course of the central axis of the bone. See C of Figure 1.
From the bone’s proximal to distal extremes, the central axis was divided
into deciles. See 0.1 to 0.9 of

From the central axis at each decile, the perpendicular distance to the lateral border of the bone was measured in tenths of millimetres. The same was determined to the medial border of the bone.
For each bone, for each decile proximal to distal, all the lateral distances were tabulated in order followed by all the medial distances. These scores approximated the bone contours and when treated by the correlational r they took on the attributes of proportionalities thereby permitting comparisons regardless of image magnification. The correlational r expressed the mathematical magnitude of the similarity from 0.0, no similarity, to 1.0, perfect in similarity.
This test was validated by determining the correlation of the bone with itself at a different magnification. This test should be independent of image magnification and the correlation should approach 1.0.
This test also was validated by determining the correlation of the same bone in two different atlases. Given atlases with perfect fidelity, this test ought to be independent of the atlas employed and the correlation ought to approach 1.0.
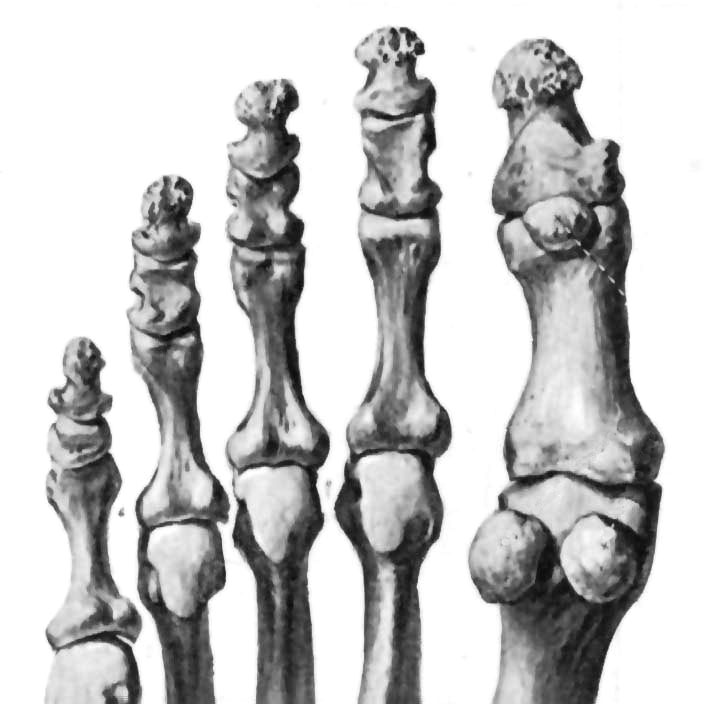
The criterion for similarity was that a correlation exceed the correlation between the phalanx 1 toe bone and its anatomical neighbour, the phalanx 2 toe bone. A correlation equal to or less than that was considered as dissimilar as a bone for an anatomically neighbouring bone. A correlation equal to or less than the one between the phalanx 1 and phalanx 3 toe bones was considered as grossly dissimilar as a bone for an anatomical neighbour two bones away. See Figure 2.


The Ardipithecus ramidus kadabba AME-VP-1/71 bone was correlated with the baboon (Papio cynocephalus) bone (see Figure 3), the chimpanzee (Pan troglodytes) bone (see Figure 4), and the human (Homo sapiens) bone (see Figures 5 and 6). The baboon, chimpanzee and human bones also were compared to each other.
Results
The plantar view of bone AME-VP-1/71 as shown in Nature correlated 0.975 with the same bone at 2.26 greater magnification. That suggested that this test was independent of any magnification or reduction in the photographic images of the fossil. The photograph of AME-VP-1/71 shown in Time correlated 0.983 with the same image at double the magnification. This result confirmed that magnification did not significantly affect the correlation and additionally demonstrated that the test was reliable and calibrated for the effects of magnification.
The dorsal view of the fourth digit, phalanx 1, human toe bone shown in Swindler and Wood correlated 0.906 with the same human bone in Spalteholz.5 That suggested that the atlas employed would lower the correlation to about 0.9. However, the plantar views correlated 0.764 and that lack of agreement between views seemed worthy of further investigation. To illumine the cause of the disparity, plantar and dorsal views were correlated for each atlas. For the Spalteholz atlas, the plantar and dorsal views correlated 0.936 whereas for the Swindler and Wood atlas their plantar and dorsal views correlated 0.605. That suggested that the cause of the lower correlation between atlases for the plantar views was the lower reliability of the plantar view in the Swindler and Wood atlas compared to the Spalteholz atlas. This result suggested that the atlas employed lowered the correlation to about 0.906 for the reliable views.
The phalanx 1 toe bone was correlated 0.657 with the phalanx 2 toe bone and 0.271 with the phalanx 3 toe bone.5 For this study, a correlation of 0.657 or less was considered dissimilar and a correlation of 0.271 or less grossly dissimilar. See Figure 2.
The AME-VP-1/71 bone shown in Nature was from a left foot but it was compared to the same bone in the right foot because the atlases did not show the left foot. The baboon, chimpanzee and human S&W correlations were made with the Swindler and Wood atlas whereas the human S correlations were made with the Spalteholz atlas. Accordingly, the plantar view of AME-VP-1/71 correlated 0.575 with the baboon bone, 0.491 with the chimpanzee bone, 0.743 with the human S&W bone, 0.613 with the human S bone. The baboon and chimpanzee bones were correlated 0.856. These results suggested that the AME-VP-1/71 bone had some similarity to humans, less to baboons and least similarity to chimpanzees. However, all of those similarities were lower than the baboon-chimpanzee similarity. The lack of higher correlations with humans and chimpanzees and the chimpanzee correlation out of the evolutionary order suggested further research.
Accordingly, another analysis was done with a more distinct plantar image of the AME-VP-1/71 bone.6 In this analysis the AME-VP-1/71 bone was correlated 0.615 with the baboon bone, 0.461 with the chimpanzee bone, 0.761 with the human S&W bone, 0.681 with the human S bone. Each correlation was slightly higher than the previous analysis with the chimpanzee correlation slightly lower. The lack of evolutionary sense, the suggestion that chimpanzees evolved to baboons, persisted and none of these correlations were as high as the 0.856 baboon-chimpanzee correlation.
This suggested investigating the dorsal view of AME-VP-1/71 and correlating it to the dorsal views in the atlases. The AME-VP-1/71 bone correlated 0.460 with the baboon bone, 0.308 with the chimpanzee bone, 0.739 with the human S&W bone, 0.873 with the human S bone. Except for human S bone, the correlations were lower and in the same order. The baboon-chimpanzee correlation was 0.806. This analysis yielded no additional clarification.
In a final attempt at clarification, for each bone the dorsal view data were combined with the plantar view data. For these combined data, the AME-VP-1/71 bone correlated 0.475 with the baboon bone, 0.377 with the chimpanzee bone, 0.729 with the human S&W bone, 0.722 with the human S bone. The baboon-chimpanzee correlation was 0.820 whereas the baboon-human correlations were 0.329 and 0.549 for S&W and S, respectively. The chimpanzee-human correlations were 0.197 and 0.332 for S&W and S, respectively. These combined data confirmed that the AME-VP-1/71 bone had scant similarity to humans (compare Figure 1 with Figure 2, Phalanx I), was dissimilar to baboons and most dissimilar to chimpanzees. The monkey was similar to the ape, dissimilar to the human and the most dissimilar was the ape and human. The ancestral sequence suggested was not the Haile-Selassie and Robinson monkey-great ape-human but rather great ape-monkey. These objective analyses identified the Haile-Selassie and Robinson conclusions as farfetched speculations.
Discussion
Oxnard did complex multivariate statistical analyses and found, ‘Nor, however, can man be described as a mosaic of other forms. In almost all studies man lies quite separately from the spectra of non-human species ….’ This multi-dimensional method, employing rotations by means of matrix algebra and a computer, found essentially what the present simpler method found, but the complex method had no cutoff for similarity, which the present simpler method has.7 This may also give insight to the concern that a simpler two-dimensional method may fall short on three dimensional objects. Like errors, shortcomings do not help but rather hinder finding differences and high correlations. If a simpler method finds what a complex method finds, then that suggests that all the complexity may not be necessary. This should also clarify whether or not confounding variables like dimorphism may need special consideration. If dimorphism were critical, then it would act like error or anything else causing undue variability and would diminish correlations toward zero rather than allow a correlation as high as 0.906.
From the 18 scores, the present method approximates the outline of the bone. When those scores are treated by the correlational r, they acquire the attributes of proportionalities. If the same bone is magnified, the proportions remain constant and that is why the r’s for the two independent magnification trials were so high and so close, 0.975 and 0.983. The bone and its magnified image have the same contours, and the high correlation indicates a high degree of similarity. Subjectively, investigators may be confused by size but the r is not. Had there been no measurement error, the r’s would have been 1.0, perfect in similarity.
If confounding variables and much error are inherent in the atlas materials, then the correlations would tend toward zero while losing the power to discriminate. Contrarily, if the atlases were the product of careful work, then the correlations should be high and should reliably discriminate. And that is what was found. Human bone in one atlas compared to another correlated 0.906 and that for this study was considered high and reliable. The same bone at different magnifications was 0.975. When tried again with a different image it was 0.983. For this study, that demonstrated that the test was reliable, it determined consistently, and was calibrated for the effects of magnification. The test thereby proved valid, reliable and calibrated, and anyone should be able to reproduce those findings, which means it is objective as well. That contrasts sharply with the subjective, unquantifiable method of inspection employed by Haile-Selassie which likely is unreliable and uncalibrated.
Errors tend to be normally and stochastically distributed about the true value. For tests of statistical significance, they increase the variability which is in the denominator of say a t-test thereby giving a ‘t’ too small to be significant. In the case of a correlation, the tendency to scatter equally above and below the true value yields an r close to 0.00. Therefore, errors or shortcomings are likely to hinder finding statistical significance or a high correlation. If statistical significance is found, or if a high correlation is found, it is in spite of errors or confounding variables not because of them.
Dimorphism or gender is a confounding variable and would have an effect similar to anything else causing error or variability. It hinders finding significance or a high correlation. For both atlases, the human sample number and gender(s) were unknown. For the Swindler and Wood atlas, there were six chimpanzees and 22 baboons all of unspecified genders. If genders whether in small or large samples made a significant difference, then the humans probably would not have correlated as high as 0.906 and the baboons and chimpanzees probably would not have correlated across species lines as high as 0.820.
A photograph may be taken of a bone, and even if not to scale, its proportionalities will be the same as the original. From a scale photograph, a tracing may be made and the same may be said of it. From several preparations, their tracings may be overlaid and the median may be identified and used in an atlas. If all of the same bones had their shapes quantified and averaged, then the rendering from those digital data would be very close to the one graphically determined. The graphical method may be similar to the methods used for atlases and if it were quite faulty then an r of 0.906 should not have been possible. Perhaps anatomists deserve more respect than they are usually accorded.
Some investigators assume that atlases may be constructed with artistic license rather than carefully from dissections. That was not the case for the atlases used in this study. Swindler and Wood stated in their preface, ‘The illustrations of Papio and Pan have been drawn from original dissections of twenty-two baboons and six chimpanzees, and represent composite illustrations based upon the specimens.’4 Spalteholz stated in the author’s preface, ‘The illustrations, in all cases, have been faithfully drawn from original preparations, but at the same time no copy of a definite individual case, but always a composite from several sections has been made.’5

The human bones used by Spalteholz in 1900 probably were different from the human bones used by Swindler and Wood in 1973, yet for the reliable views the r was 0.906. Let us pretend that the Spalteholz bones were the ancestors of the Swindler and Wood bones. Let us suppose further that a human and his ancestor is likely to be correlated about 0.906. Let us call this the top of the scale of similarity. Now, what is needed is a lower limit of similarity. Any lay person will observe the significant difference in proportions between phalanx 1 and phalanx 2 and would easily judge them dissimilar (see Figure 2). That level of dissimilarity had an r of 0.657. Now, there is objectively established a range of similarity expected to be between 0.906 and 0.657. If r is lower than 0.657, then the bones are as dissimilar as a bone for an anatomically different bone. The expectation is that if there is ancestry, then the bones will be in the range of 0.657 to 0.906. This was confirmed with humans compared to humans, 0.906, and across species lines monkeys compared to apes, 0.820. Monkeys and humans, even more so apes and humans, were as dissimilar as for an anatomically different bone. The method established those criteria objectively, validly, reliably and with calibration. Scientifically, that may be considered a higher standard than the subjective inspection that led to the extravagant claims of Haile-Selassie and Robinson.
This study used the atlases as standards and they worked well. Had the atlases been faulty, then the correlations would have approached zero in every comparison and the study would have failed. Had that happened, then there may have been the need to go out and obtain materials. The human bones and those of chimpanzees and baboons would have been available for random sampling but not the solitary fossil bone. Therefore, the sampling approach would have a benefit only if the atlases were unreliable and if the sampling would have provided more reliable data. Given reliable atlases, it did not seem a judicious expenditure of resources to duplicate work already adequate for successful studies.
A case may be made for postponing the study and the report until an adequate sample of fossils is found. That is like allowing what is perceived to be the only statistical design available, sample statistics, to dictate scientific activity. Statistics are meant to be a tool, not the master. The alternative is to seek or invent a statistical design that accommodates a sample of one. By making repeated determinations on that sample of one, which may be considered random samples normally distributed, then conventional statistical tools may be applied. The statistical universe is that sample of one and all conclusions apply only to that sample of one. That is exactly what was wanted for the present study of a sample of one fossil. Now the question of scientific acceptability arises. To my knowledge this sample of one method was first reported in the Research Quarterly in 1959 and was again used successfully and reported in a different protocol in 1967. It is ideally suited to individualized pilot protocols especially those requiring stringent, multiple controls. It probably has been used and reported in the literature many times since 1967 and has been accepted undisputed for generalized application by the scientific community for more than 40 years. That was the concept used in this study and that is why the conclusions apply to that fossil as a sample of one and the specified atlases each treated as if they were samples of one.
Haile-Selassie must consider that what he observes as dorsal tilting may not be objectively and quantitatively established in the life-long bipedal range. Even if it were in the life-long bipedal range, it is not likely to have only that similarity and no other. It is not likely to pass a subjective test but fail an objective test. If it did, then which is to be doubted? The protocol for an experiment must be repeatable by any scientist in any laboratory and yield the same answer. Anyone may obtain the results of the present study. Very few have access to the fossil and although accessible, a better image than that shown in Nature was not made available. By contrast, the present method is available worldwide. Haile-Selassie’s is confined to a locked drawer as was done with the Piltdown forgery of 1912–1952. The verdict ought to be put in the hands of scientists worldwide. Although available and upon request, Haile-Selassie would not even allow out a better image than the indistinct one that appeared in Nature.
Haile-Selassie apparently is using his eyes and subjective judgement on the slant of a joint surface, a joint surface unavailable to the scientific community. Were it available, a way might be found to do a parallel, objective, valid, reliable, calibrated test of similarity. It is not available, so scientists denied access must do the best they can with whatever is available and that is what was done. It is unfortunate that those with access do not take seriously their responsibility to the scientific community at large and devise the objective tests themselves. That they do not, that they make much from such meagre data, that they deny access to even a better image, and that the results of this modest study are so contradictory make one wonder whether their purpose is to avoid rather than welcome scientific scrutiny, acceptance and application. Was the intent instead to proceed directly from a letter to the editor of Nature1 to the acclaim from a cover story in Time magazine?2 If they are confident of their work with nothing to hide, then they should behave like bona fide scientists and not like those involved in the Piltdown forgery.3
Summary and conclusions
In summary, the results of this objective statistical study suggest that the AME-VP-1/71 bone had scant similarity to human bone, was dissimilar to baboon bone and was most dissimilar to chimpanzee bone. The baboon bone was similar to the chimpanzee and dissimilar to human bone. The chimpanzee was most dissimilar to humans. Human bone had no similarity to monkey or ape bone. Therefore, these objective ancestry analyses for fossil bones suggest that the conclusion of Haile-Selassie and Robinson, that Ardipithecus ramidus kadabba was an ancestor of apes and humans that walked on two legs, is farfetched speculation.
References
- Haile-Selassie, Y., Late Miocene hominids from the Middle Awash, Ethiopia, Nature 412:178–181, 2001. Return to text.
- Robinson, S., Paleontology, one giant step for mankind, Time 158(3):54–61, 2001. Return to text.
- Walsh, J.E., Unraveling Piltdown: The Science Fraud of the Century and Its Solution, Random House, New York, pp. 124–125, 1996. Return to text.
- Swindler, D.R. and Wood, C.D., An Atlas of Primate Gross Anatomy: Baboon, Chimpanzee, and Man, University of Washington Press, Seattle, pp. 52–55, 1973. Return to text.
- Spalteholz, W., Hand Atlas of Human Anatomy, Volume I, S. Hirzel, Leipzig, pp. 146–149, 1900. Return to text.
- Sarfati, J., Time’s alleged ‘ape-man’ trips up (again)! Journal of Creation 15(3):7–9, 2001. Return to text.
- Oxnard,C.E., The place of australopithecines in human evolution: grounds for doubt? Nature 258:389–395, 1975. Return to text.

Readers’ comments
Comments are automatically closed 14 days after publication.