

Journal of Creation 23(1):94–98, April 2009
Browse our latest digital issue Subscribe
Karyotypic and allelic diversity within the canid baramin (Canidae)
Previous studies suggest that all dog-like creatures (canids, family Canidae) belong to a single created kind. As unclean animals, all modern canids are descendants of two canids preserved on the Ark during the Flood. This pair of canids would have carried a limited amount of genetic diversity. They would be expected to have had a fairly uniform arrangement of chromosomes (low karyotypic diversity) and up to four different versions of any particular gene (allelic diversity). Today there is considerably more karyotypic and allelic diversity within the canids. The patterns imply that more than random mutation and natural selection are involved; instead, certain genetic components appear designed to change and numerous designed mechanisms may be involved in driving many of these changes. This suggests that God, in His infinite wisdom, designed animals to be able to undergo certain genetic mutations which would enable them to adapt to a wide range of environmental challenges while minimizing risk.

There is a need to more fully describe intrabaraminic (within kind) variation on a genetic level for understanding the basis for the variety we see within baramins today. It has been pointed out that the majority of mutations are near neutral.1 Yet intuitively, I would expect random (chance) ‘errors’ in such a complex system to be more consistently disastrous unless the system was designed to change.2 If genetic systems were designed to allow for such changes, then mutations (changes in the nucleotide sequence of DNA) are not necessarily just ‘errors’ or ‘accidents’. On the contrary, some mutations may be directed to allow animals to adapt in the present fallen world. By examining intrabaraminic genetic diversity, we should be able to discover a clearer picture regarding the role of mutations in the development of the diversity found in animals today.
Previous baraminic studies have identified all canids (family Canidae) as belonging to a single baramin.3 Since they are unclean animals, all living canids would have descended from a single breeding pair preserved on the Ark about 4,500 years ago.4,5 This historical information is important because it suggests there was a limited amount of diversity present in canids at that time. Today, this family is represented by 34 species that are widely distributed around the world.6 There are considerable data available on the karyotypic and allelic diversity in protein coding genes for several of these species. A brief overview of the data is presented here.
Karyotype
The family Canidae exhibits the most highly rearranged karyotypes* of any family within the order Carnivora. Normal diploid numbers vary from 34 for the red fox (Vulpes vulpes) to 78 for the domestic dog (Canis familiaris) and dhole (aka Asiatic Wild Dog; Cuon alpinus) (table 1). The Arctic fox (Alopex lagopus) is polymorphic for a centric fusion; diploid numbers of 49 and 48 are found in individuals carrying one or two copies respectively of this fusion. Phylogenomic analysis suggests that 82 may have been the ancestral karyotype. Within the 10 species that have been studied in detail it appears that approximately 80 rearrangements have occurred. This includes numerous fusions, both centric and tandem, fissions, pericentric inversions and/or centromere transpositions.7 Several paracentric inversions, and even whole arm (telomere to centromere) inversions, have been implicated based on the differences in loci order among species (figure 1).8,9

Evidence of similar rearrangements is present within other baramins and even within some species.10–12 Detailed studies of rearrangements in ruminants strongly suggest that numerous designed mechanisms operate to repair breaks, silence an extra centromere, adjust amounts of heterochromatin and possibly alter the position of the centromere.13 The fact that such rearrangements often become fixed within a species suggests that they may be beneficial under certain circumstances. However, fixing these rearrangements also likely required a small population, since it is difficult to fix even beneficial mutations in a large population.14 Thus, rearrangements should not be viewed as a major genetic accident from which animals occasionally may recover. Instead, the presence of multiple designed mechanisms enabling translocations to occur while maintaining viability of the animal suggests that such rearrangements are likely helpful for adaptation in the present fallen world. This is not to say that such rearrangements are without risk. For example, many heterozygous carriers experience some decline in fertility. Occasionally there are more serious results with infertility and/or serious chromosomal aberrations in the offspring.13 Furthermore, these types of rearrangements certainly don’t explain the origin of chromosomes.
The red fox and both subspecies of raccoon dog carry B chromosomes as part of their normal karyotype.7 These small, supernumerary chromosomes can vary in number both within as well as among individuals. Generally their numbers are low, with three to five being typical for the red fox.15 They usually contain significant amounts of repetitive sequences and, until recently, it was thought that they did not contain any protein coding genes. However, the canid B chromosomes have been found to contain the KIT gene, which encodes a transmembrane tyrosine kinase receptor involved in the proliferation, migration and differentiation of hematopoietic, melanoblast, and primordial germ cells. Adjacent sequences were detected, including the RPL23A pseudogene and, in the raccoon dog only, a portion of the more distal KDR gene. This suggests that the B chromosomes were derived from an autosome in a common ancestor and have been lost in other lineages descending from this ancestor. Further studies need to be done to determine if the KIT gene of B chromosomes is actually transcribed.16
Major histocompatibility complex genes
The major histocompatibility complex (MHC) consists of a number of genes involved in immune function and which are known for high allelic diversity. Several dog leukocyte antigen (DLA) genes have been evaluated for polymorphisms. As of 2006, there were 90 alleles recognized for DLA-DRB1, 22 for DLA-DQA1 and 54 for DLA-DQB1, with more expected to be discovered.17 High levels of polymorphism are generally considered a sign of a healthy population, although some dog breeds and wild mammals have low MHC diversity with no apparent ill effects. The DLA genes are on dog chromosome (CFA) 12.18 Some DLA haplotypes are associated with various canine autoimmune diseases such as primary immune mediated hemolytic anemia, polyarthritis, hypothyroidism and diabetes.19 However, it is important to recognize that these haplotypes do not cause disease directly; instead, they may be risk factors that affect the likelihood of disease development. As suggested previously, there is risk in maintaining sufficient variability to adapt in the present fallen world.
Dopamine receptor D4 gene
There are two portions of the dopamine receptor D4 (DRD4) gene that are variable in dogs. The first is in exon 1 where the two known alleles differ by a 24-base pair (bp) indel.20 Interestingly, humans also are polymorphic in this region with a 12-bp duplication and a 13-bp deletion having been identified.21 The latter is particularly intriguing as it is found in 2% of the human population and is not associated with any known disease; yet the frameshift is predicted to result in a truncated, non-functional protein.22

The second polymorphic region is found in exon 3. There are eight alleles that have been identified in dogs.20 A number of these have been identified in wolves. The alleles differ by variable number tandem repeats (VNTRs) of 12-and 39-bp (figure 2). A similar pattern has been observed in humans, where a 48-bp segment is repeated from 2 to 10 times. These variations are believed to influence behaviour because certain alleles have been shown to be associated with the novelty-seeking personality trait in humans, primates and dogs.23 VNTRs have been identified in exon 3 of the DRD4 gene of nearly all mammals examined except rodents. The length of the repeated segments varies among taxa, but is consistently a multiple of three.24
This bias of indels, particularly VNTRs, in base pairs that are multiples of three does not appear to be explicable by natural selection. If essentially random, approximately one-third of indels should be multiples of three unless a frameshift, which often results in a premature stop codon and a nonfunctional protein, is lethal or significantly detrimental. It does not appear that frameshifts in DRD4 would be subject to such selection pressure, since a frameshift mutation is carried by a number of normal humans and knock-out mice.20,22 Furthermore, variability in this gene appears to contribute to variability in personality. The number of alleles in canids (greater than eight, as the raccoon dog has a separate allele identified25) is greater than the maximum of four alleles expected in the pair of canids on the Ark. Humans also carry more alleles than can be attributed to Adam and Eve. This suggests that this gene was designed to vary in a rather unusual way to enhance variability in personality and perhaps other traits as well.
Olfactory genes
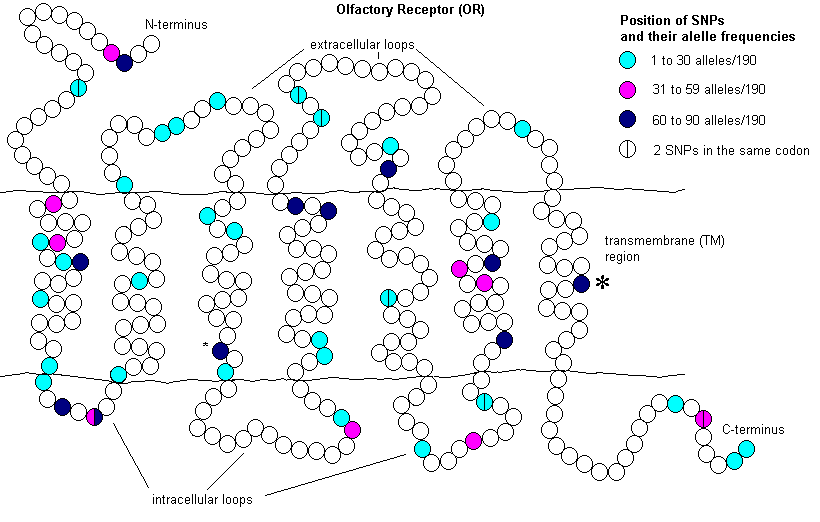
Olfactory (smell) receptor (OR) genes are seven transmembrane receptors. While 1,094 OR genes have been identified in the dog,26 the canine repertoire of odorant molecules is significantly greater than this. This appears to be from a complex combinatorial code. Odorant molecules can bind 20 or more ORs depending on their concentration. ORs can bind more than one odorant molecule. Through interpretation of the complex signalling patterns, dogs are able to detect an incredibly wide array of individual odorants and a large number of mixtures.27
In one study, 16 OR genes were examined in 95 dogs from 20 different breeds. All genes were polymorphic ranging from two to 11 alleles per gene. There was an average of one change per 920 sequenced nucleotides, which is much higher than most coding sequences and a random sampling of non-coding sequences. Of the 98 single nucleotide polymorphisms (SNPs) identified, 55 resulted in an amino acid change and 30 of these involved changes to a different amino acid group. These changes were found throughout the protein (figure 3), mostly in variable or highly variable regions within OR genes. However, two come from highly conserved regions, one in transmembrane (TM) 3 and the other in TM7.28
Five of the 16 genes had an allele with a disrupted open reading frame. These were from one of the four indels identified or an SNP introducing a stop codon. Pseudogenization of OR genes is fairly common. In poodles, 18% of ORs are pseudogenes while 20.3% (or 222/1094) are in the boxer. Interestingly, 17 of the OR pseudogenes in the poodle were not found in the boxer, and 22 of those found in the boxer were not found in the poodle.28
It may be premature to assume there is no purpose in mutation or pseudogenization within OR genes.29 There is a tremendous amount of redundancy in OR genes which may have been designed to allow for future specialization. For example, a study involving Drosophila sechellia, a highly specialized vinegar fly that feeds solely on fruit from Morinda citrifolia, a shrub which strongly repels related species of flies, suggests that pseudogenization of ORs and gustatory (taste) receptors has occurred nearly 10 times faster than in the closely related species D. simulans. For those genes which remained intact, D. sechellia appears to have fixed non-synonymous substitutions at a consistently higher rate than synonymous substitutions compared to the same genes in D. simulans.30 Therefore, the ability of OR genes to be modified or pseudogenized may be an important design element introduced by the Creator.
Conclusion
The two canids preserved on the Ark would be expected to have carried a fairly uniform karyotype and up to four alleles for non-duplicated genes. This brief examination of present-day karyotypes and several groups of genes indicates that significant diversity has arisen since the Flood. Several different lines of evidence suggest that many of these mutations may have some benefit to the animal. For example, intrabaraminic chromosomal comparisons have implicated numerous designed mechanisms which control chromosomal changes in a way that maintains viability of the animal. The fact that such mechanisms appear to be operating suggests there is purpose to chromosomal rearrangements. The fact that different karyotypes often are fixed in different species within a baramin seems to support this concept as well.
The various genes examined here appear to handle mutations very well. In fact, it is generally believed that the high allelic diversity in the MHC genes is important for a healthy population. The redundancy in ORs and the pattern of mutation and pseudogenization in these genes suggests that these genes were designed to vary so that animals can adapt to different environments. Finally, the striking non-random pattern of VNTR mutations, all in lengths divisible by three, when there is no known selection that could produce this non-random pattern, strongly suggests that in some instances there are designed mechanisms driving mutations. The patterns seen here suggest that God, in His infinite wisdom, designed animals to be able to undergo genetic mutations which would enable them to adapt to a wide range of environmental challenges while minimizing risk.
Glossary | |
|---|---|
| Autosome: | a chromosome that is not a sex (X or Y) chromosome. |
| Centric fusion: | combining of two acrocentric (centromere near one end) chromosomes to form a new chromosome with the centromeres adjacent to each other. See figure 1. |
| Centromere transposition: | a change in the position of the centromere on the chromosome without a change in gene order. This rearrangement can be very difficult to distinguish from a pericentric inversion. |
| Frameshift: | an insertion or deletion (indel) that shifts the three-base-pair reading frame of the gene. A frameshift will often result in loss of function of the protein. |
| Haplotype: | a region of DNA usually inherited together; a group of alleles that are closely linked. |
| Heterochromatin: | sections of DNA containing highly repetitive sequences and few genes. Despite appearing inactive, these regions are important for proper function. The amount of heterochromatin appears to be adjusted following chromosomal rearrangements. |
| Karyotype: | the appearance of the chromosomes within an individual at metaphase, the time during cell division when the chromosomes are clearly seen. |
| Open reading frame: | the portion of DNA that is read (copied into RNA) and may be used for protein formation. |
| Knock-out mice: | mice in which the specific gene under study is disabled (knocked out). Studies with knock-out mice have been very helpful in determining the function of genes. |
| Paracentric inversion: | an inversion in one chromosome arm that does not include the centromere. See figure 1. |
| Pericentric inversion: | an inversion in a chromosome that includes the centromere. See Figure 1. |
| Phylogenomic: | comparison of the genomes of organisms within a group to attempt to reconstruct ancestry. |
| Single nucleotide polymorphism (SNP): | a difference in a single base in the DNA sequence; a change in which a single base pair differs from the usual base pair in that position. |
| Tandem fusion: | combining of two chromosomes where the end of one chromosome attaches to the end or centromeric region of another chromosome. See figure 1. |
| Tandem repeats: | multiple copies of the same base sequence on a chromosome. See figure 2. |
References
- Sanford, J.C., Genetic Entropy & The Mystery of the Genome, Elim Publishing, New York, 2005. Return to text.
- See Lightner, J.K., Designed to inhabit the earth, A review of The Plausibility of Life: Resolving Darwin’s Dilemma, Journal of Creation 22(1):33–36, 2008, and Williams, A., Facilitated variation: A new paradigm emerges in biology, Journal of Creation 22(1):85–92, 2008. Return to text.
- Siegler, H.R., The magnificence of kinds as demonstrated by canids, CRSQ 11(2):94–97, 1974. See also Wood, T.C., The current status of baraminology, CRSQ 43(3):149–158, 2006. Return to text.
- Genesis 7:1–10; Leviticus 11:27 Return to text.
- Ussher, J., The Annals of the World, Translated and edited by L. and M. Pierce, Master Books, Green Forest, AR, p. 19, 2003. Return to text.
- Fahey, B. and Myers, P., ‘Canidae’ (Online), Animal Diversity Web, 2000, animaldiversity.ummz.umich.edu/site/accounts/information/Canidae.html, accessed August 30, 2008. Return to text.
- Graphodatsky, A.S. et al., Phylogenomics of the dog and fox family (Canidae, Carnivora) revealed by chromosome painting, Chromosome Research 16(1):129–143, 2008. Return to text.
- Szczerbal, I. et al., Development of a cytogenetic map for the Chinese raccoon dog (Nyctereutes procyonoides procyonoides) and the arctic fox (Alopex lagopus) genomes, using canine-derived microsatellite probes, Cytogenetic and Genome Research 102: 267–271, 2003. Return to text.
- Szczerbal, I. et al., FISH mapping of 10 canine BAC clones harbouring genes and microsatellites in the arctic fox and the Chinese raccoon dog genomes, Journal of Animal Breeding and Genetics 123(5):337–342, 2006. Return to text.
- Lightner, J.K., Identification of species within the sheep-goat kind (Tsoan monobaramin), Journal of Creation 20(3):61–65, 2006. Return to text.
- Nachman, M.W. and Myers P., Exceptional chromosomal mutations in a rodent population are not strongly underdominant, Proceedings of the National Academy of Sciences of the United States of America 86(17):6666–6670, 1989. Return to text.
- Lightner, J.K., Changing chromosome numbers, Journal of Creation 20(3):14–15, 2006. Return to text.
- Lightner, J.K., Karyotype variability within the cattle monobaramin, Answers Research Journal 1:77–88, 2008. Return to text.
- ReMine, W., More precise calculations of the cost of substitution, CRSQ 43(2):111–120, 2006. Return to text.
- Rausch, V.R. and Rausch, R.L., Karyotype of the Red Fox, Vulpes vulpes L., in Alaska, Northwest Science 53(1):54–57, 1979. Return to text.
- Yudkin, D.V. et al., Mapping of KIT adjacent sequences on canid autosomes and B chromosomes, Cytogenetic and Genome Research 116:100–103, 2007. Return to text.
- Kennedy, L.J. et al., Association of a common dog leukocyte antigen class II haplotype with canine primary immune-mediated haemolytic anaemia, Tissue Antigens 68:502–508, 2006. Return to text.
- Angles, J.M. et al., Frequency and distribution of alleles of canine MHC-II DLA-DQB1, DLA-DQA1 and DLA-DRB1 in 25 representative American Kennel Club breeds, Tissue Antigens 66:173–184, 2005. Return to text.
- Kennedy et al., ref. 17. Return to text.
- Ito, H. et al., Allele frequency distribution of the canine dopamine receptor D4 gene exon III and I in 23 breeds, Journal of Veterinary Medical Science 66(7):815–820, 2004. Return to text.
- Hejjas, K. et al., Novel repeat polymorphisms of the dopaminergic neurotransmitter genes among dogs and wolves, Mammalian Genome 18(12):871–879, 2007. Return to text.
- Nöthen, M.M. et al., Human dopamine D4 receptor gene: frequent occurrence of a null allele and observation of homozygosity, Human Molecular Genetics 3(12):2207–2212, 1994. Return to text.
- Hejjas, K. et al., Association of polymorphisms in the dopamine D4 receptor gene and the activity-impulsivity endophenoyte in dogs, Animal Genetics 38(6):629–633, 2007. Return to text.
- Larsen, S.A. et al., Identification and characterization of tandem repeats in exon III of dopamine receptor D4 (DRD4) genes from different mammalian species, DNA Cell Biol 24(12):795–804, 2005. Return to text.
- Inoue-Murayama, M. et al., Sequence Comparison of the Dopamine Receptor D4 Exon III Repetitive Region in Several Species of the Order Carnivora, Journal of Veterinary Medical Science 64(8): 747–749, 2002. Return to text.
- Quignon, P. et al., The dog and rat olfactory repertoires, Genome Biology 6(10):R83, 2005. Return to text.
- Benbernou, N. et al., Functional analysis of a subset of canine olfactory receptor genes, Journal of Heredity 98(5):500–505, 2007. Return to text.
- Tacher, S. et al., Olfactory receptor sequence polymorphism within and between breeds of dogs, Journal of Heredity 96(7):812–816, 2005. Return to text.
- This is not only true because some pseudogenes have previously been shown to have important non-protein coding function, but also because a human OR pseudogene containing multiple premature stop codons has recently been demonstrated to code for a functional protein. Lai, P.C. et al., An olfactory receptor pseudogene whose function emerged in humans: a case study in the evolution of structure-function in GPCRs, Journal of Structural and Functional Genomics, September 19, 2008. Return to text.
- McBride, C.S., Rapid evolution of smell and taste receptor genes during host specialization in Drosophia sechellia, Proceedings of the National Academy of Sciences of the United States of America 104(12):4996–5001, 2007. Return to text.




Readers’ comments
Comments are automatically closed 14 days after publication.