

Journal of Creation 28(3):45–47, December 2014
Browse our latest digital issue Subscribe
On pterosaurs, and flights of the evolutionary imagination
Pterosaurs From Deep Time by David M. Unwin
Pi Press, 2006

The author is, or was, curator of fossil reptiles and birds at the Museum of Natural History, Humboldt University, Berlin. Pterosaurs were a group of flying reptiles that, according to the evolutionary-uniformitarian geologic timescale, flourished during the Mesozoic (figure 1). Though contemporaneous with the dinosaurs, they were not related to them (so the popular term ‘flying dinosaurs’ is wrong). Even though they probably resembled other volant vertebrates (figure 2), pterosaurs were built differently from birds or bats.
This work is relatively nontechnical. It contains numerous visual aids and summaries of information, including a stratigraphic column of pterosaur occurrences, a series of phylogenies, an atlas of pterosaur occurrences, a table of localities and pterosaur-bearing formations, and a list of currently recognized pterosaur species and higher-level taxons to which they belong.
Pterosaur biology
This work features a lucid description of the major aspects of pterosaur paleobiology. One learns that pterosaurs ranged from those with a 20 cm wing-span to giants with 10 m wingspans.
Unwin touches on the challenges of recognizing fossil species. In the past, juveniles and adults of the same species had mistakenly been assigned to separate species (the same happened with dinosaurs1). So have the male and female of the same species. This can be generalized. As more fossils are collected, there is a tendency to lump previously named species into one.
This work includes an impressive amount of detail on evidence that pterosaurs were excellent flyers. Adaptations for flying included hollow bones, limb morphology, fusion of vertebrae, specialized wrists, and the pteroid bone that supported an aileron-like skin flap.
Pterosaurs had a form of body hair. The function or functions that it served are speculative. Hair may have served as a form of insulation. It also may have created a layer of air, next to the body, that would reduce drag during flight. If the hair had different colours, this could have served diverse functions, ranging from camouflage to attracting mates.
Unwin suggests that pterosaurs were warm-blooded, even if not to the degree of modern birds and mammals. The need for sustained flight would have required a thermal physiology that would allow for a sustained energy output that is unlike that of typical reptilian physiology.
The author also discusses technical advances in paleontology and paleobiology that have been applied to pterosaurs. One of these is the use of computers and sophisticated cameras. Colour filters are used and these help bring out features on the pterosaur fossils that are not visible under ordinary light.
Evolution deified
In Unwin’s thinking, evolution is much more than a purported scientific explanation for the existence of living things. It is nothing less than an all-powerful transcendental process. He comments:
“Ask almost anybody: What is the most powerful thing in the world? And they will probably reply: ‘gravity’, or ‘love’, or ‘money’. All these answers and many others, including ‘Microsoft’, are wrong, because the right answer is ‘evolution’. Every single living thing, from the smallest microbe to the biggest blue whale, is a consequence of evolution, even the most complex thing in the known universe—our brains. Evolution also built pterosaurs” (p. 90).
Those who contend that ‘religion’ (meaning God and Creation) and ‘science’ (meaning evolution) belong to non-overlapping magisteria are missing the point, if not being disingenuous. It is obvious that evolution has, at least to some evolutionists, clearly assumed God-like and therefore God-replacing properties. Worse yet, evolution has a hard time explaining anything, let alone everything. This is notably true of pterosaurs, as elaborated below.
‘Ghost lineages’—evolution’s failures
One striking feature of this work is the inferred evolutionary deployment of the pterosaurs, shown by Unwin in his figure 10.2 (p. 228). The striking feature of this figure is the frank portrayal of ‘missing’ pterosaurs, that is ‘ghost lineages’. Figure 1 is a simplified and schematized portrayal of Unwin’s figure 10.2.
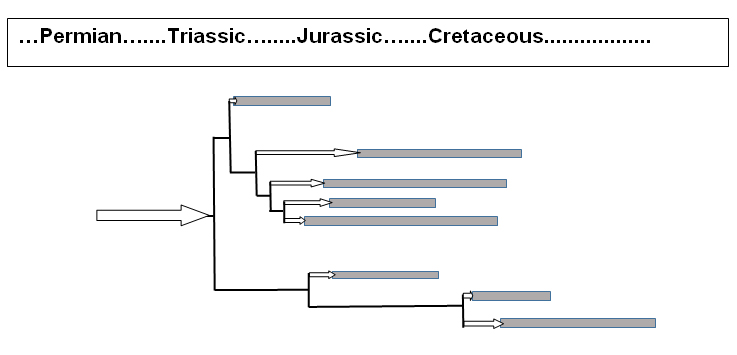
There are a number of implications of these ad hoc ghost lineages. Let us first consider stratomorphic intermediates, which are related to the common evolutionary argument that the order of appearance of organisms in the fossil record agrees with the order of evolutionary branching as inferred by morphology (cladistics analyses). This often is not so and every ghost lineage (arrow in figure 1) is a tacit admission of a glaring conflict between the two. The organism in question does not appear in the ‘correct’ position in the stratigraphic record. The ghost lineage is thus an imaginary extension of the organism’s fossil record to where it ‘should’ appear in order to agree with the predictions of cladistic analysis.
Second, the ghost lineage can be an indirect admission that the organism is morphologically discontinuous with its closest evolutionary relatives. For this reason, a significant interval of time (the ‘ghost lineage’) must be invented in order to give it time to evolve all the differences it has from its closest relatives (as inferred from cladistics relationships).
The standard evolutionary explanation (or rationalization) for ghost lineages is an incomplete fossil record. In the case of pterosaurs, this is buttressed by the contention that pterosaurs, having thin skeletons suitable for flying, fossilize poorly. This argument is less than convincing. To begin with, pterosaur fossils, though uncommon in comparison with marine invertebrate fossils, are not exactly exotic rarities. Unwin notes that 5,000 to 6,000 pterosaur individuals have been collected so far (p. 51). Nor are there only a handful of places on Earth where pterosaurs have been found. His tabulation of pterosaur-bearing locations (his figure 3.9, pp. 52–53) is telling. There are 38 such locations, and that on every continent except Antarctica. Moreover, 21 of these 38 locations have yielded at least 10 pterosaur specimens, and seven of these locations have yielded at least 100 pterosaur fossils. Therefore, as usual, the excuse of an incomplete fossil record ends up begging the question: the only ‘evidence’ of the incompleteness is the lack of transitional forms, the very thing the ‘incompleteness’ is supposed to explain.
Evolutionary assumptions are imposed upon the evidence
Interpretations of the fossil record always begin with an a priori acceptance of organic evolution. No matter what turns up in the fossil record, some evolutionary scenario can always be weaved around it. This is as true of modern cladistically-based reconstructions as it was with the early inferences of ancestor-descendant relationships. As an example of all this, consider the following account by Unwin:

“Establishing how different species of pterosaur were related to one another was for many years primarily based on their overall degree of similarity and their geological age. Because the Early Jurassic prow-jawed pterosaur Dorygnathus appeared to be quite similar to the Late Jurassic prow-jawed Rhamphorynchus, it was not only assumed that they were more closely related to each other than to any other pterosaur, but that Rhamphorhynchus was directly descended from Dorygnathus. … Pterosaurs have recently been subjected to several ‘phylogenetic analysis’ among the results of which is the discovery that while Dorygnathus and Rhamphorhynchus still belong in the same prow-jawed clan—the rhamphorynchines—they have been joined by two or three relatives and are no longer thought to be directly related to one another” (p. 61).
Systematic discontinuities in the evolution of pterosaurs
The author Unwin tacitly admits the conjectural nature of pterosaur evolution:
“As figure 4:3 illustrates, paleontologists don’t really know where this group should sit within the diapsid family tree. The reason for this confusion is simple—a complete lack of protopterosaurs that might link this group to other diapsids” (p. 64).
“As you will doubtless recall from Chapter 4, the exact point at which the pterosaurs branched off from other diapsid reptiles have yet to poke their heads out of the fossil record (or if they have done so, they are keeping their identity well-hidden)” (p. 230).
“Such a degree of variety among the earliest known pterosaurs tells us that, evolutionarily speaking, a lot must have happened to the group prior to the mid-Late Triassic” (p. 233).
Unwin then contrasts two hypotheses: the ‘big bang’ and the ‘iceberg’. According to the former, the pterosaurs underwent a geologic rapid evolutionary deployment just before their first appearance in the mid-Late Triassic. However, according to the ‘iceberg’ model (referring to the proverbial ‘tip of the iceberg’), the pterosaurs originated long before their first appearance in the fossil record—perhaps even in the Permian (corresponding to the large arrow in figure 1).
Furthermore, there are huge gaps between different kinds of pterosaur. The usual theory postulates that the ‘primitive’ pterosaurs like Rhamphorynchus had long tails and needed to stabilize flight at the cost of manoeuverability. Supposedly, the more evolved or ‘derived’ pterosaurs had large brains, thus enabling more precise control to stabilize flight, and therefore they did not need a long tail, and so were more stable. The ‘more evolved’ members of the order Pterosauria are the suborder Pterodactyloidea (the term ‘pterodactyl’ should be confined to the suborder but is popularly misused to mean pterosaurs in general). However, as Unwin admits:
“Pterodactyloids’ appearance in the Late Jurassic is almost as dramatic as the debut of pterosaurs in the Triassic. Frustratingly, we still have no evidence of their ancestors—intermediate forms somewhere between a rhamphorynchoid and pterodactyloid—but this is nothing to get excited about. Such ‘missing links’ are notoriously rare. What is more surprising is the almost complete absence of any fossil remains of pterodactyloids in the first 50 million years of the Jurassic, even though the genealogical map illustrated in figure 10.2 suggests that they had already appeared by the end of the Early Jurassic and quite possibly well before” (pp. 244–245). [For illustration, see the bottom-most three taxons in figure 1].
The nearest creature that could be an intermediate is Darwinopterus, but this ruins the evolutionary story—it had the advanced brain and the long tail together. Unwin himself admitted elsewhere how unexpected this was:
“But the strange thing about Darwinopterus is that it has a head and neck just like that of advanced pterosaurs, while the rest of the skeleton, including a very long tail, is identical to that of primitive forms.”2
Conclusion
The pterosaurs were a group of fascinating flying reptiles. They were probably excellent flyers. Pterosaur evolution, however, is especially unconvincing. Numerous ad hoc ghost lineages have to be invented to cover up the discrepancies between the pterosaur fossil record and the predictions of evolutionary theory.
References and notes
- Handwerk, B., A third of dinosaur species never existed?, National Geographic News, 9 October 2009. Return to text.
- Unwin, D., cited in McGrath, M., New flying reptile fossils found, news.bbc.co.uk, 14 October 2009. Return to text.







Readers’ comments
Comments are automatically closed 14 days after publication.