
Species were designed to change, part 1
But how much change is allowed?

Creationists do not, and have never, taught that God created all species as we see them today. People in the past certainly believed in ‘the fixity of species’ but this was more due to the pervasive influence of the pagan Greek philosopher Aristotle than to anything in the Bible. One of Darwin’s primary influences, the lawyer-turned-geologist Charles Lyell, taught fixity of species. He also believed that species were placed in “centres of creation”. In other words, species were more or less created in their current locations. Many of Darwin’s points in the Origin of Species and later books were designed to refute Lyellian and Aristotelian arguments, although we can see their influences in his earlier writings.
The biblical Creation/Flood/Dispersion model really has nothing to do with Darwin’s objections. He was not arguing against biblical teaching. In fact, there is an excellent example in the Bible itself that shows how fast species can change: the episode where Jacob bred spotted and speckled animals from all-white flocks (Genesis 30). Clearly, species are not fixed. The Bible talks about a worldwide dispersion of the animals on the Ark after the Flood. Thus, in no way does the Bible suggest that all species were created where they are found today. Darwin observed that many species on the Galápagos islands were clearly related to those on the South American mainland, but they were subtly different as well. This refuted Lyell’s “centres of creation” model, yet the observations are even better explained by post-Flood migration paths (which I will cover in Part 2 of this series).
Creatures were designed to change
The prevailing idea within creationist circles is that God created distinct “kinds” that were front-loaded with a large amount of genetic diversity that could then be used as the kinds turned into our modern species. Dr. Peer Terborg used the term “pluripotent baranomes”. Pluripotent describes something that can turn into many other things. A pluripotent stem cell, for example, can be used to grow any organ in the body. A baranome is a play on words. The suffix “–ome” is a made-up root word used by biologists to denote a total collection of objects. Therefore, a genome is a collection of all the ‘genes’ found in an individual or species, the proteome is all the proteins found in an individual or species, and the microbiome is the total collection of microbes in a human or animal body. Bara is the Hebrew word for “create”. Therefore, in the same way that a genome represents all the genes in a single person, a baranome represents all the genomes in a single baramin (that is, a created kind). These were filled with reams of useful information, so the original horses, whatever they may have looked like, could turn into all the varieties we see both today and in the fossil record.
When God designed life, He knew how far livings things could be pushed. In effect, he wrote a tolerance code into the design specs of each created kind. Think of the great variety we see in the various kinds. Horses, for example, range from very small to very large. They can be shaggy or short-haired. Their colours range from white to red to brown to black, with many potential combinations in the same animals. Much (but not all) of this was coded into the first horses.
Initial conditions allow for front-loaded information
Some of this initial information was in the form of heterozygous alleles (i.e. genetic variation). We are not told how many individuals in each kind God created, except for humans. Given an initial population, God could have engineered any amount of genetic diversity into the genomes of the created kinds. Even humans could have had more initial diversity than is normally carried in two people because there was nothing stopping God from engineering a different genome into every one of Adam and Eve’s reproductive cells.1 Alternatively, Adam and Eve could have been created with normal reproductive cells but with a lot more genetic diversity than modern humans currently have.2 In either case, the amount of diversity we see among humans today would depend, in large part, on how many children they had.

Some of that initial information could also have been in latent form. God did not have to hard-code every single future trait ever seen in every species but could have allowed for the possibility of certain changes to take place. For example, retrotransposons (‘jumping genes’) can turn genes on or off depending on where they are located in the genome. Entire new phenotypes could arise, depending on where these genetic elements are, or are not, located in the genome. No “new” information needs to be created if this was part of the original design criteria for life. Yet, “new” characteristics (‘phenotypes’) could certainly arise.
The four-dimensional structure of the genome also plays a role in how the genome can change over time. Retroviruses like HIV insert themselves into the genome. Yes, they change the genome. However, this is not a random process. They are much more likely to integrate in areas that are open, not areas that are deeply buried in coils of DNA. They are also more likely to integrate in areas that affect the structure of DNA itself.3 The structure of DNA also influences the timing, rate, and types of mutation (discussed below) that will happen in the genome. Thus, when God programmed those linear strings of DNA we call ‘the genome’, He knew how it would fold up into a 3-D shape and how that 3-D shape could be modified in the 4th dimension (i.e. time). Part of this high-level organization involved forward planning. By putting a gene in a specific place, God was setting up specific possibilities for future modifications.
Another way to create new phenotypes is through recombination. Chromosomes get scrambled each generation. This brings genetic variants together in new and unique ways. Let’s say that two genes that affect height and muscularity can be found near each other in the genome of some species. We’ll call the first one T, for tall. We’ll call the second one M, for muscular. Let’s say they each have a recessive form. We’ll call them t (short) and m (thin) and that God originally set it up so that any one chromosome had either T-M or t-m (figure 1).

Using dogs as an example, since individuals carry two copies of each chromosome, they could be:
- TTMM (tall and muscular, like a mastiff)
- TtMm (also tall and muscular, since t and m are recessive)
- ttmm (short and thin, like a Chihuahua)
But note what is lacking. No individual can be TTmm or ttMM. It is not possible to get an animal that is tall and thin (like a greyhound) or short and muscular (like a bulldog). OK, but what if a recombination occurred between the two genes (figure 2)?
Suddenly, we have two new combinations (figure 3). We started with only three possibilities. We now have five (TTMM, TTmm, TtMm, ttMM, or ttmm). Given that each species has thousands of genes, recombination could be a continual source of new phenotypes, especially early on.

Yet another way to create new phenotypes is through gene gain and gene loss. Genetics has thrown us a few curve balls over the years. Take the lowly E. coli bacterium. It has a genome of 4.6 million letters and contains 4,288 protein-coding genes. Only that’s not really true, for different bacteria in that same species can carry different sets of genes. If something they need is being produced by another species living nearby, for example, deleting that gene will be beneficial. Why spend time and effort maintaining that section of DNA and making proteins when the thing you need is free? Because of this, members of the same species carry different genes depending on the environment in which they are living. If you add up all the genes carried by all E. coli in the world, you end up with what we now call a pangenome, with up to 5,500 genes. The pangenome concept gives God a way to create a phenomenal amount of potential diversity. The difference in gene content from one organism to another in the same kind could have been pre-engineered or could have arisen through mutation.
Mutations: engineered probability
Clearly, God could engineer a lot of potential changes into creation. However, there is something else we have to consider: mutations. God was not naïve about mutations. When He wrote out each genome, He knew how that code could be mutated. In effect, part of the design specifications of life included an understanding of how DNA would be modified over time.
Yet, God also knew that unchecked mutation would not be a good thing, because most mutations are harmful, simply because there are many more ways to break something than to make it. Therefore, He also created multiple amazing, complex, and well-engineered error-checking and repair systems. There is a different repair system for each type of mutation, from single-letter misincorporations to near-fatal double strand breaks.
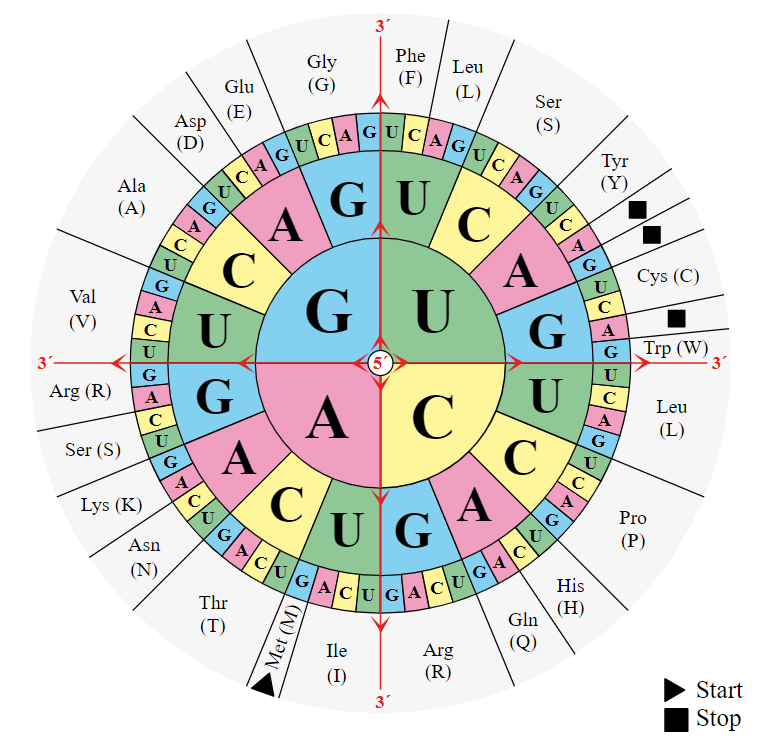
Even so, some mutations still slip through this system, so He added yet another level of complexity. You may have heard that three letters of DNA codes for one amino acid in a protein. Because there are four bases, there are 64 possible 3-letter codons (43) but only 20 amino acids, meaning some amino acids are coded by more than one codon (figure 4). But what they usually forget to tell you in school is that the ‘redundancy’ in the amino acid code is optimized to slow the rate of change. There are millions of ways to assign 20 amino acids to 64 codons, but God picked one in which the genetic code helps to minimize the effect of mutation. When a mutation does occur, the amino acid is often unchanged, and when an amino acid is changed, it is often changed to a similar amino acid. Most other arrangements are not nearly as effective at reducing the potential effects of mutation.
If a mutation has little to no effect, it is effectively neutral. There is no problem with thinking this and so we can take a clue from the evolutionary community, many of whom believe that most mutations are harmless, because some certainly are. Yet, mutations are not truly random. You see, some mutations are much more likely than others. Thus, given any initial starting point, any genetic system will change in a preferred direction. This is completely contrary to the evolutionary idea, where all possibilities must stay on the table. Instead, we can see specific types of changes building up in different things, like the high number of C to T and G to A mutations seen in the human H1N1 influenza virus since 1917 (figure 5).4 We documented a 1% change in the number of As and Gs and about a 0.5% change in the number of Us and Cs between 1917 and 2009. This was due to the simple fact that not all mutations are equally likely, because some bases are more thermodynamically stable than others. When God wrote out the DNA sequences of the original baranomes, He certainly knew the direction in which they would change. This was all part of His initial plan.

To that end, some genes are more protected than others, with robust and continual error checking. Other genes are much more likely to mutate than others. For example, the genes that control melanin production in mammals are highly mutable. There are also examples of genetic systems that seems to be designed to rapidly change in order to exploit new resources. An example of this would be the ability of certain bacteria to be able to digest nylon, a man-made fiber never seen in nature. Instead of being an example of evolution, these bacteria clearly have a designed ability to adapt. In fact, geneticists are moving away from the old, now outdated, idea that mutations are inherently random.5 Non-randomness could be due to differences in the chemical code (i.e. the C to T change is much more common than T to C, and some genes are enriched in C), the position within the nucleus when the genome is in its normal 3D configuration, the presence of duplicate copies of a gene, the presence of repetitive DNA close by, the near presence of non-B-DNA (non-B (including cruciform loops, Z-DNA, and quadruplex looping), and many other factors. God knew this when he wrote the initial code. He knew.
Mutation is not a surprise to God. He is the one who wrote the laws of chemistry, and He knew how oxygen and water would continually react with DNA when He chose to use DNA as the information storage mechanism in the cell. He designed the DNA repair systems and engineered a known fault tolerance level into them. He knew that the DNA polymerases that copy DNA would occasionally make mistakes and He designed them with this error rate in mind. If He wanted more faithful replication, He would have designed the system more stringently. If He wanted sloppy DNA copying, this would have been easy to do. Again, He knew.
Another factor that God deliberately designed into living things is something called epigenetics. In our cells, there are all sorts of switches that can turn genes on and off. Genes that are not needed can be turned off. The cell can stick extra carbon atoms on the DNA in that region (a process called methylation) or it can modify the histone proteins the DNA wraps around. Unlike mutations, many of which don’t have any measurable effect, epigenetic changes are designed to produce phenotypic change. Yet, this depends on a specific set of letters being in place. If those letters mutate, the epigenetic switch can be broken, either in the ‘off’ position or the ‘on’ position.
Of course, most of this applies after the Fall. There is some small scope for mutations prior to sin and death entering the world, but these would have to be carefully orchestrated by God. In the same way that Adam would never have been able to trip over a root and plummet headlong off a cliff before the Curse, mutations would never have been able to create deformities in living things. But this does not mean that there was no chance for any change in any letter in any baranome in all of creation.
Natural selection
As organisms within each kind reproduce, it is natural to think that some will be better fit for some environments while others would be better suited to other environments. This is the essence of natural selection. Much has been written about this already, and it is not a threat to the creation model. In fact, in some ways natural selection would even apply to the time before the Fall (see Natural Selection in Paradise). After the Fall, however, selection would go into overdrive. As the environment, especially after the Flood, changed, the phenotype of specific organisms would be very important. Death also reigned. Survival became an all-important factor as organisms now struggled with environmental conditions that were never seen before (like the Ice Age). It is good to remember that natural selection is not a creative force. It is, however, something that happens automatically as organisms reproduce over time. This is one of the many factors that combine to create change over time. Notice that it was also saved for last. Darwin once thought natural selection was the preeminent force driving change over time. Instead, it is a minor component among a diverse array of factors that God put into his creation.
Conclusions
God is the master engineer. He did not just create life; He designed life to robustly respond to the environment. He overdesigned living things so that they could withstand, so far, thousands of years of mutational accumulation. His designs were holistic, in that He accounted for what each kind would need in the future. He built redundancy in the genomes to ward off the effects of decay, but He also designed specific aspects of the genome to change via mutation.
In part 2 of this article, we will deal with population-level changes, speciation events, the rate of speciation, and how much “change” life can tolerate. But let’s leave off with a verse that tells of the greatness of our Creator, Romans 11:33–36:
“Oh, the depth of the riches and wisdom and knowledge of God! How unsearchable are his judgments and how inscrutable his ways!
“For who has known the mind of the Lord,
or who has been his counselor?”
“Or who has given a gift to him
that he might be repaid?”
For from him and through him and to him are all things. To him be glory forever. Amen.”
References and notes
- Sanford, J., Carter, R., Brewer, W., Baumgardner, J., Potter, B., and Potter, J., Adam and Eve, designed diversity, and allele frequencies. In Proc. 8th Eighth International Conference on Creationism, ed. J.H. Whitmore, pp. 200–216, 2018; digitalcommons.cedarville.edu/icc_proceedings/vol8/iss1/8. Return to text.
- Carter, R.W. and Powell, M., The genetic effects of the population bottleneck associated with the Genesis Flood, J. Creation 30(2):102–111, 2018. Return to text.
- Cereseto, A. and Giacca, M. Integration site selection by retroviruses. AIDS Rev. 6(1):13–21, 2004. Return to text.
- Carter, R.W., More evidence for the reality of genetic entropy, J. Creation 28(1):16–17, 2014. Return to text.
- Cooper, D.N., et al., On the sequence-directed nature of human gene mutation: the role of genomic architecture and the local DNA sequence environment in mediating gene mutations underlying human inherited disease. Hum. Mutat. 32(10):1075–1099, 2011. See also Evolution’s well-kept secret: Mutations are not random! Return to text.










Readers’ comments
Comments are automatically closed 14 days after publication.