
Table of Contents

Unit 1
Argument: Creationism is religion, not science
Argument: Evolution is compatible with Christian religion
Argument: Evolution is true science, not ‘just a theory’
Unit 2
Argument: Natural selection leads to speciation
Argument: Some mutations are beneficial
Argument: Common design points to common ancestry
Argument: ‘Bad design’ is evidence of leftovers from evolution
Argument: The fossil record supports evolution
Unit 3
Argument: Probability of evolution
Argument: ‘Irreducible complexity’
Argument: Evolution of sex
Argument: Evolution of mankind
Common arguments for evolution that have been rejected
Common arguments for creation that should not be used
Refuting Evolution 2—Chapter 10
A sequel to Refuting Evolution that refutes the latest arguments to support evolution (as presented by PBS and Scientific American).
Argument: ‘Irreducible complexity’
Evolutionists say, ‘Examples of supposed “irreducible complexity” (such as the eye, the complex cell and the flagellum) can be explained.’
First published in Refuting Evolution 2, Chapter 10
This chapter will examine how evolutionists respond to the ‘irreducible complexity’ argument in three areas: the eye, the complex cell and the flagellum. Scientific American states the problem this way:
14. Living things have fantastically intricate features—at the anatomical, cellular and molecular level— that could not function if they were any less complex or sophisticated. The only prudent conclusion is that they are the products of intelligent design, not evolution.
This ‘argument from design’ is the backbone of most recent attacks on evolution, but it is also one of the oldest. In 1802, theologian William Paley wrote that if one finds a pocket watch in a field, the most reasonable conclusion is that someone dropped it, not that natural forces created it there. By analogy, Paley argued, the complex structures of living things must be the handiwork of direct, divine invention. Darwin wrote On the Origin of Species as an answer to Paley: he explained how natural forces of selection, acting on inherited features, could gradually shape the evolution of ornate organic structures. [SA 83]
Indeed, Gould, who was an expert on the history of evolution, agreed that Darwin was writing to counter Paley. This is another way of saying that he had an anti-theistic agenda,1 as discussed in chapter 2. This doesn’t stop many churchian academics kowtowing to every pronouncement made by Darwin and his God-hating successors, who in return regard them as contemptuously as Lenin regarded his ‘useful idiot’ allies in the West.2
Could the eye have evolved?
It’s interesting to note that the eye, which evolutionists claim is an example of ‘bad design’ leftover from evolution (previous chapter), presents their greatest challenge as an example of superb ‘irreducible complexity’ in God’s creation. Scientific American says:
Generations of creationists have tried to counter Darwin by citing the example of the eye as a structure that could not have evolved. The eye’s ability to provide vision depends on the perfect arrangement of its parts, these critics say. Natural selection could thus never favor the transitional forms needed during the eye’s evolution—what good is half an eye? Anticipating this criticism, Darwin suggested that even ‘incomplete’ eyes might confer benefits (such as helping creatures orient toward light) and thereby survive for further evolutionary refinement. [SA 83]
First, this overlooks the incredible complexity of even the simplest light-sensitive spot. Second, it’s fallacious to argue that 51 percent vision would necessarily have a strong enough selective advantage over 50 percent to overcome the effects of genetic drift’s tendency to eliminate even beneficial mutations.3
Biology has vindicated Darwin: researchers have identified primitive eyes and light-sensing organs throughout the animal kingdom and have even tracked the evolutionary history of eyes through comparative genetics. (It now appears that in various families of organisms, eyes have evolved independently.) [SA 83]
Scientific American contradicts itself here. If the evolutionary history of eyes has been tracked through comparative genetics, how is it that eyes have supposedly evolved independently? Actually, evolutionists recognize that eyes must have arisen independently at least 30 times because there is no evolutionary pattern to explain the origin of eyes from a common ancestor. What this really means is that since eyes cannot be related by common ancestor, and since they are here, and only materialistic explanations are allowed, hey presto, there’s proof that they evolved independently!
Simulation of eye evolution
PBS 1 goes to great lengths to convince us that the eye could easily have evolved. Dan Nilsson explained a simplistic computer simulation he published in a widely publicized paper.4 Taking his cue from Darwin, who started with a light-sensitive spot when ‘explaining’ the origin of the eye, Nilsson’s simulation starts with a light-sensitive layer, with a transparent coating in front and a light-absorbing layer behind.
Here is how the simulation proceeds. Firstly, the light-sensitive layer bends gradually into a cup, so it can tell the direction of light rays increasingly well. This continues until it is curved into a hemisphere filled with the transparent substance. Secondly, bringing the ends together, closing the aperture, gradually increases the sharpness of the image, as a pinhole camera does, because a smaller hole cuts out light. But because of the diffraction of light if the hole is too small, there is a limit to this process. So thirdly, the shape and refractive index gradient of the transparent cover change gradually to a finely focusing lens. Even if we were generous and presumed that such computer simulations really have anything to do with the real world of biochemistry, there are more serious problems.
However, the biochemist Michael Behe has shown that even a ‘simple’ light-sensitive spot requires a dazzling array of biochemicals in the right place and time to function. He states that each of its ‘cells makes the complexity of a motorcycle or television set look paltry in comparison’ and describes a small part of what’s involved:5
When light first strikes the retina a photon interacts with a molecule called 11-cis-retinal, which rearranges within picoseconds to trans-retinal. (A picosecond [10-12 sec] is about the time it takes light to travel the breadth of a single human hair.) The change in the shape of the retinal molecule forces a change in the shape of the protein, rhodopsin, to which the retinal is tightly bound. The protein’s metamorphosis alters its behavior. Now called metarhodopsin II, the protein sticks to another protein, called transducin. Before bumping into metarhodopsin II, transducin had tightly bound a small molecule called GDP. But when transducin interacts with metarhodopsin II, the GDP falls off, and a molecule called GTP binds to transducin. (GTP is closely related to, but different from, GDP.)
GTP-transducin-metarhodopsin II now binds to a protein called phosphodiesterase, located in the inner membrane of the cell. When attached to metarhodopsin II and its entourage, the phosphodiesterase acquires the chemical ability to ‘cut’ a molecule called cGMP (a chemical relative of both GDP and GTP). Initially there are a lot of cGMP molecules in the cell, but the phosphodiesterase lowers its concentration, just as a pulled plug lowers the water level in a bathtub.
A transparent layer is also far more difficult to obtain than the researchers think. The best explanation for the cornea’s transparency is diffraction theory, which shows that light is not scattered if the refractive index doesn’t vary over distances more than half the wavelength of light. This in turn requires a certain very finely organized structure of the corneal fibers, which in turn requires complicated chemical pumps to make sure there is exactly the right water content.6
Therefore, these simulations do not start from simple beginnings but presuppose vast complexity even to begin with. Also, in their original paper, the researchers admitted ‘an eye makes little sense on its own,’ because the ability to perceive light is meaningless unless the organism has sophisticated computational machinery to make use of this information. For example, it must have the ability to translate ‘attenuation of photon intensity’ to ‘a shadow of a predator is responsible’ to ‘I must take evasive measures,’ and be able to act on this information for it to have any selective value. Similarly, the first curving, with its slight ability to detect the direction of light, would only work if the creature had the appropriate ‘software’ to interpret this. Perceiving actual images is more complicated still. And having the right hardware and software may not be enough—people who have their sight restored after years of blindness take some time to learn to see properly. It should be noted that much information processing occurs in the retina before the signal reaches the brain.
It is also fallacious to point to a series of more complex eyes in nature, and then argue that this presents an evolutionary sequence. This is like arranging a number of different types of aircraft in order of complexity, then claiming that the simple aircraft evolved into complex ones, as opposed to being designed. For one thing, eyes can’t descend from other eyes per se; rather, organisms pass on genes for eyes to their descendants. This is important when considering the nautilus eye, a pinhole camera. This cannot possibly be an ancestor of the vertebrate lens/camera eye, because the nautilus as a whole is not an ancestor of the vertebrates, even according to the evolutionists!
Rotary motors in the bacterial flagellum
Scientific American cites another difficult example of irreducible complexity—the rotary motors on bacterial flagellum, but it really has no answers.
15. Recent discoveries prove that even at the microscopic level life has a quality of complexity that could not have come about through evolution.
‘Irreducible complexity’ is the battle cry of Michael J. Behe of Lehigh University, author of Darwin’s Black Box: The Biochemical Challenge to Evolution. As a household example of irreducible complexity, Behe chooses the mousetrap—a machine that could not function if any of its pieces were missing and whose pieces have no value except as parts of the whole.
What is true of the mousetrap, he says, is even truer of the bacterial flagellum, a whiplike cellular organelle used for propulsion that operates like an outboard motor. The proteins that make up a flagellum are uncannily arranged into motor components, a universal joint, and other structures like those that a human engineer might specify. The possibility that this intricate array could have arisen through evolutionary modification is virtually nil, Behe argues, and that bespeaks intelligent design. [SA 84]
Indeed, it does (see diagram below).
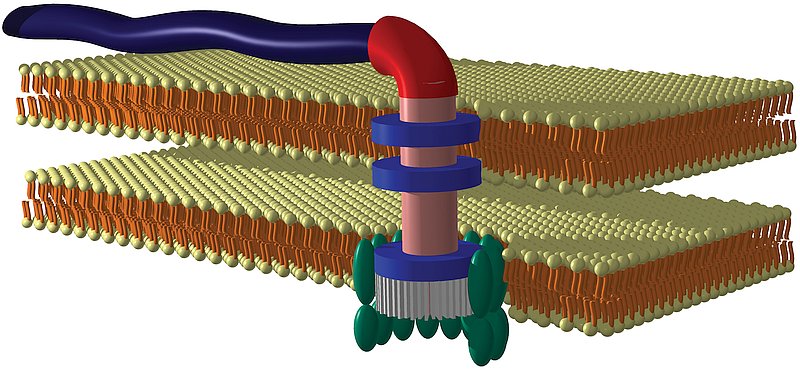
Bacterial flagellum with rotary motor, with the following features:
- Self assembly and repair
- Water-cooled rotary engine
- Proton motive force drive system
- Forward and reverse gears
- Operating speeds of up to 100,000 rpm
- Direction reversing capability within 1/4 of a turn
- Hard-wired signal transduction system with short-term memory
[from Bacterial Flagella: Paradigm for Design, video,
www.arn.org/news-videos/videos.html]
He makes similar points about the blood’s clotting mechanism and other molecular systems.
Yet evolutionary biologists have answers to these objections. First, there exist flagellae with forms simpler than the one that Behe cites, so it is not necessary for all those components to be present for a flagellum to work. The sophisticated components of this flagellum all have precedents elsewhere in nature, as described by Kenneth R. Miller of Brown University and others. [SA 84]
Miller is hardly the epitome of reliability. Behe has also responded to critics such as Miller.7
In fact, the entire flagellum assembly is extremely similar to an organelle that Yersinia pestis, the bubonic plague bacterium, uses to inject toxins into cells. [SA 84]
This actually comes from the National Center for Science Education’s misuses of the research of Dr Scott Minnich, a geneticist and associate professor of microbiology at the University of Idaho. He is a world-class expert on the flagellum who says that belief in design has given him many research insights. His research shows that the flagellum won’t form above 37°C, and instead some secretory organelles form from the same set of genes. But this secretory apparatus, as well as the plague bacterium’s drilling apparatus, are a degeneration from the flagellum, which Minnich says came first although it is more complex.8
The key is that the flagellum’s component structures, which Behe suggests have no value apart from their role in propulsion, can serve multiple functions that would have helped favor their evolution. [SA 84]
Actually, what Behe says he means by irreducible complexity is that the flagellum could not work without about 40 protein components all organized in the right way. Scientific American’s argument is like claiming that if the components of an electric motor already exist in an electrical shop, they could assemble by themselves into a working motor. However, the right organization is just as important as the right components.
The final evolution of the flagellum might then have involved only the novel recombination of sophisticated parts that initially evolved for other purposes. [SA 84]
Minnich points out that only about 10 of the 40 components can be explained by co-option, but the other 30 are brand new. Also, the very process of assembly in the right sequence requires other regulatory machines, so is in itself irreducibly complex.9
Blood clotting
Scientific American cites another serious problem for evolution—blood clotting.
Similarly, the blood-clotting system seems to involve the modification and elaboration of proteins that were originally used in digestion, according to studies by Russell F. Doolittle of the University of California at San Diego. So some of the complexity that Behe calls proof of intelligent design is not irreducible at all. [SA 84]
This is once more a lot of bluff by the atheist Doolittle, or at least poor reading comprehension. He cited recent experiments showing that mice could survive with two of the components of the blood clotting cascade (plasminogen and fibrinogen) eliminated. This supposedly showed that the current cascade was not irreducibly complex but clearly reducibly complex. But the experiment really showed that the mice lacking both components were better off than one lacking only plasminogen, because the latter suffer from uncleared clots. But the former are hardly as healthy as Doolittle implied, because the only reason they don’t suffer from uncleared clots is that they have no functional clotting system at all! A non-functioning clotting system (despite possessing all the many remaining components) is hardly an evolutionary intermediate that natural selection could refine to produce a proper clotting system. Rather, this experiment is evidence against this, because the next step (i.e., from lacking both plasminogen and fibrinogen to fibrinogen only) would be selected against because of the uncleared clots.10
Complexity of a different kind—‘specified complexity’—is the cornerstone of the intelligent-design arguments of William A. Dembski of Baylor University in his books The Design Inference and No Free Lunch. Essentially, his argument is that living things are complex in a way that undirected, random processes could never produce. The only logical conclusion, Dembski asserts, in an echo of Paley 200 years ago, is that some superhuman intelligence created and shaped life.
Dembski’s argument contains several holes. It is wrong to insinuate that the field of explanations consists only of random processes or designing intelligences. Researchers into nonlinear systems and cellular automata at the Santa Fe Institute and elsewhere have demonstrated that simple, undirected processes can yield extraordinarily complex patterns. Some of the complexity seen in organisms may therefore emerge through natural phenomena that we as yet barely understand. But that is far different from saying that the complexity could not have arisen naturally. [SA 84]
Talk about blind faith! But in practice, as Dembski points out, specified complexity in all cases but biology is used as evidence of design, including the search for extraterrestrial intelligence. Since biological complexity is the only exception proposed by evolutionists, it smacks of special pleading.11
In addition to the human eye, the flagellum, and blood clotting, there’s a host of other examples of irreducible complexity in nature. Earlier I alluded to the dynamic sticking mechanism in the legs of insects. The sticky feet of geckos is another clear example of God’s ingenuity.12 Its structure is described by its evolutionary discoverers as ‘beyond the limits of human technology.’13 Still other examples of design include the lobster eyes with their unique square reflecting geometry that inspired advanced x-ray telescopes and beam producers,14 the ATP synthase motor.
References and notes
- Wieland, C., Darwin’s real message: have you missed it? Creation 14(4):16–19, September 1992; J. Sarfati, review of K. Birkett, The Essence of Darwinism; see Evangelical compromise misses the essentials. Return to text.
- Sarfati, J., The Skeptics and their ‘Churchian’ allies. Return to text.
- See my discussion about the evolution of the eye in Stumbling over the impossible: refutation of Climbing Mt Improbable, Journal of Creation 12(1):29–34, 1998; see Eye evolution, a case study. Return to text.
- Nilsson, D.E. and Pelger, S., A pessimistic estimate of the time required for an eye to evolve, Proc. R. Soc. Lond. B 256:53–58, 1994. Return to text.
- Behe, M.J., Darwin’s Black Box: The Biochemical Challenge to Evolution, The Free Press, New York, NY, 1996, p. 46. Return to text.
- Gurney, P.W.V., Dawkins’ eye revisited, Journal of Creation 15(3):92–99, 2001. Return to text.
- Behe responds to various critics, www.trueorigin.org/behe08.asp. Return to text.
- See Scott Minnich, Bacterial flagella: spinning tails of complexity and co-option, www.idurc.org/yale-minnich.html, available via web.archive.org. Return to text.
- Unlocking the Mystery of Life, video, Illustra Media, 2002. Return to text.
- For more information, see Behe’s In defense of the irreducibility of the blood clotting cascade www.trueorigin.org/behe03.asp. Return to text.
- Grigg, R., A brief history of design, Creation 22(2):50–53, March 2000. Return to text.
- JSarfati, J., Great gecko glue? Creation 23(1):54–55, December 2000. Return to text.
- Autumn, K., et al., Adhesive force of a single gecko foot hair, Nature 405(6787): 681–685, 8 June 2000; perspective by H. Gee, Gripping feat, same issue, p. 631. Return to text.
- Sarfati, J., Lobster eyes—brilliant geometric design, Creation 23(3)12–13, June 2001. Return to text.
Note about citations: Quotations from the Scientific American article by John Rennie will be labeled ‘SA,’ followed by the page number. Quotations from, and other mentions of, the PBS-TV series ‘Evolution,’ will be labeled ‘PBS,’ followed by the episode number, e.g. ‘PBS 6’ refers to Episode 6. Return to article.
Readers’ comments
Comments are automatically closed 14 days after publication.