

Journal of Creation 8(1):105–116, April 1994
Browse our latest digital issue Subscribe
Homo erectus ‘to’ modern man: evolution or human variability?
Abstract
An interesting change is taking place in creationist circles in respect of the status of the taxon Homo erectus and its relationship to Homo sapiens sapiens. This development is paralleled by a similar change of direction in evolutionary thinking, and in both cases it seems likely that the impetus is being largely propelled by the discovery of the erectus specimen KNM-WT 15000 in Africa in 1984. This attitudinal shift has connotations for the whole topic of alleged evolution of human beings. In this brief paper it is proposed to track these amended attitudes and the implications for the creation-evolution controversy.
Introduction
With the discovery of Java and Peking Homo erectus fossils (the former was previously called Pithecanthropus erectus, and Peking Man was originally named Sinanthropus pekinensis), in 1891–1893 and 1927–1937 respectively, evolutionary theory received a considerable and much-needed boost. Until Dubois’ Java discoveries, the only alleged link between man and the apes had been a few Neanderthal specimens. The Piltdown hoax of 1912–1914 was not uncovered until 1953, by which time it had played a considerable part in the early skepticism by most authorities toward the Taung-child australopithecine discovery in South Africa in 1924.
With the appearance of the Javan and Peking fossils it seemed that evolutionary theory had been vindicated to a sizeable degree, and Pithecanthropus (ape-man) became a common term in public as well as in palaeoanthropological circles.
After the Piltdown fraud was exposed, the australopithecines came into favour as a transitional form linking an ape-like common-ancestor to human beings, and this link was further strengthened by later finds of both erectus and australopithecine fossils, mainly in East and South. Africa. By the early 1970s, more finds including australopithecus-like material classified as Homo habilis, made it appear that there was now a fairly substantial chain of progressive evolution from a bipedal chimp-like ancestor right through to modern man - A. afarensis, H. habilis, H. erectus, archaic H. sapiens, Neanderthal man and finally Cro-Magnon or modern man. With the rise of the post-World War II creationist movement, largely sparked by the epic work of Whitcomb and Morris in 1961,1 one of the most urgent tasks involved was how to respond to this apparent chain of evolutionary progression. In the intervening years since then, creation-oriented scientists have made a number of attacks on the validity of most of these forms, some of them being of high technical quality, others a little less well-informed.
The central feature of this paper mostly involves the so-called erectus section of the chain, where a quite noticeable change of direction is well underway, both in the creationist and the evolutionist camps. Far from dismissing erectus forms as being only large extinct apes or frauds, the pendulum is now swinging to the view that most, if not all erectus specimens are indeed full members of the human race. With the discovery of the Turkana ‘Boy’ WT 15000 in 1984 in Kenya, it is no longer possible to hold to the position that Homo erectus was only a large-brained pongid.

In evolutionary circles it is becoming increasingly common to argue that although H. erectus forms are still on the ‘road’ from ape to human, the morphological distinctions between all human-type forms are insufficient to justify a separate species classification for erectus—that is, that all post-habiline forms (erectus, archaic and modern sapiens plus the Neanderthals), could be subsumed into a single species—H. sapiens, with a subspecific distinction at most.
(The times and ages mentioned here are those commonly used in evolutionary scientific literature and are used for ease of identification only. I do not accept an earth history of millions of years.
Australopithecines and Habilines
In a recent paper I argued at length that neither of these forms are human ancestors but are simply extinct varieties of extremely chimp-like pongids,2 and much the same view is taken by Gish,3 by Lubenow,4 and by Beasley,5 although Lubenow regards Homo habilis as an invalid taxon, being represented by a mixture of Homo sapien fossils and ape-like fossils.6 A strong creationist case exists in respect of the australopithecines and the so-called habilines, and the weight of evidence now seems to point away from any connection with humans and towards the strong likelihood that they were nothing more than extinct apes. Indeed evolution authorities Cherfas and Gribbin believe that a strong case can be made that the gracile forms are little more than varieties of extinct pygmy chimpanzees, while the robust forms are varieties of gorillas.7
Homo Erectus : Man, Ape, or Ape-man?

The Javan and Peking forms of erectus in particular came under considerable attack by creationists in the 1970s and 1980s.8,9,10 The thrust of these critiques was that all erectus forms were extremely ape-like and even possibly fraudulent. A great deal was made of the fact that almost every Peking fossil mysteriously disappeared in 1941, leaving students nothing to work on but casts (see Figure 1).11 Java Man was also regarded as suspect, on account of its discoverer Dubois having allegedly hidden two fully human Javan skulls for many years in order to strengthen his claims for the ‘Pithecanthropus’ or erectus specimen known as Java Man (see Figure 2).12 These so-called Wadjak skulls are variously reported as from nearby river gravels or from cave deposits many kilometres away. In any case, evolutionist Loring Brace claims that Dubois did make some preliminary reports about these skulls.13 Further doubt was raised because the fully modern femur of Dubois’ Java Man was found in the following year 15 metres away from where the skull cap was located.14 Brace and Montagu in 1977 state that ‘Curiously, Dubois waited until the 1920’s to also reveal he had found four more human thigh bones in the area where his Pithecanthropus material had been discovered.’ (Emphasis added.)15
(Actually the Wadjak skulls were discovered in river gravels nearby, but their age has always been a matter of some dispute.)
The Turkana ‘Boy’, KHM-WT 15000

However, with the Leakey/Walker 1984 find of the above specimen, it became clear that there was more to Homo erectus than Java and Peking Man. WT 15000 consists of almost a complete skeleton (see Figure 3).16 Only the hands and feet are missing.
The resemblances between WT 15000 and the two controversial Asian erectus forms are clear and decisive. The Turkana Boy possessed the same heavy supraorbital ridges, the same type of receding forehead, and other cranial features as Java and Peking. With an estimated age of about 11 years old at death, and a cranial capacity (EndoCranial Volume) of about 900 cc, WT 15000 is plainly a human being - even in the post-cranial features. Radiodated at about 1.6 mya (million years ago), this lad stood and walked as fully erect as do humans today.17,18 Although the brain capacity is rather small, it is still larger than some juvenile and adult humans of today.
As most of the adult cranial capacity is reached by age 10 or 11, it is likely that the adult ECV of WT 15000 would be no more than about l000–1050 cc, which is still well within the modern human range of about 800–2000 cc.19 On the same page Jue points out that a brain capacity of 1400 cc applies to the Vertesszöllos erectus specimen which is dated at around 350 kya (kiloyears ago = thousands of years ). Beasley cites Broderick who reported a measurement as low as 830 cc for a modern Wedda pygmy in Sri Lanka.20
At a height of five feet four inches or 1.6 metres, it is likely that WT 15000 was getting close to full adult height at the time of his death — this could have been anywhere between five feet five inches to maybe six feet; there is no possible way to establish final height with certainty, although a figure closer to six feet is more probable. In my high school days, I had a friend of age 15 who was short—around five feet two inches, yet by age 19 he had mushroomed to six feet. At the same age, another of my school friends was about five feet five inches yet in adulthood he had gained only another two inches. One cannot always successfully extrapolate in these matters. In all vital respects WT 15000 was as human as you or I.
Another interesting erectus specimen is skull KNM-ER 3733 dating from about 1.7 mya. It also possesses, along with other ‘ancient’ erectus forms, much the same type of cranial morphology as did Java, Peking and WT 15000, and has an ECV of approximately 850–900 cc.21,22 (See Figure 4.)23 ER 3733’ cranium is dolichocephalic, a feature also found in many Neanderthals, and it is thick, as are most erectus and Neanderthal specimens. The four erectus specimens so far mentioned are the main subject of this paper. According to Lubenow, the entire ECV range of known erectus forms runs from 700 cc for a Javan infant to 1200 cc—the largest Peking skull.24 However, the capacity of the previously-mentioned Vertesszöllos fragment from Hungary and dated at about 350 kya, is estimated at about 1400 cc, which is high for an erectus specimen.25
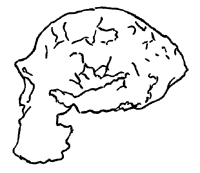
The finding of ER 3733 and WT 15000 therefore appears to strongly reinforce the validity of Java and Peking Man. The clear similarities shared by all four (where skeletal and cranial material is available), render untenable any claims that the two Asian specimens are nothing more than exceptionally large apes. Further, their affinities with both archaic sapiens and Neanderthal sapiens are so strong that it can hardly be denied that all are closely related human beings.
The question of course is - are erectus forms proof of an evolutionary progression from the apes, or are they simply temporal, regional, climatic, dietary or pathological variants of human beings?
Discussion
Naturally, one of the most important questions is that of time. If the standard geological time-scale is correct, then the very slight changes in erectus morphology over a period of 1.3 million years of existence may carry a little more weight. Yet we must also remember that this type had displayed a remarkable degree of structural stasis over the whole period from c.1.7 mya to about 350 kya. The morphology of the earliest specimens such as WT 15000 differs insignificantly from the much later specimens such as the Peking and Javan examples, the only significant difference being restricted to the endocranial volume. At full adulthood, the Turkana Boy would have possessed a capacity of about 1000–1050 cc, compared to later Chinese examples which were as high as 1200 cc.26
According to Molnar, the modern human range runs from about 700–2200 cc,27 and this puts every adult erectus specimen comfortably into the range of modern humans, and this range also covers every adult example of archaic sapiens, Neanderthal, and Cro-Magnon Man.
Writing in 1985, Pellegrino conceded that the differences between H. erectus and modern man are merely superficial.28 On the same page he even discusses the probability that H. erectus and H. sapiens are one and the same species.
Now obviously if the distinctions between erectus and modern humans are merely superficial as Pellegrino admitted, then the differences between the earliest and the latest erectus specimens, and between erectus and archaic and Neanderthal sapiens are even more superficial; that is, there is a great probability that all erectus, Neanderthal and H. sapiens are closely related, with genetic, dietary, climatic, and other environmental diversity in evidence.
As Beasley29 and Lubenow30 have recently published excellent papers on the question of the archaic H. sapiens, it is not my intention to go into that question in any great detail.
There are literally thousands of hominid fossils in existence and of these, over 300 are classified as either Neanderthal or erectus.31 We have a large enough sample to be certain about the accuracy of the diagnostic features of all groups. Admittedly taxonomic names count for less than the actual morphological structures of the various human races, past and present; but from an evolutionary viewpoint the small degree of change in erectus populations over an alleged period of one and a quarter million years must be disappointing, especially if the cranial capacities of the earliest and latest examples all lie within the modern range of humans. Pellegrino wrote in 1985: ‘(Between the first and the last erectus specimens), there are no major morphological excursions; merely a thicker brow ridge here, a subtle variation of tooth structure there, and not much else. It looks as if a substantial stretch of human evolution was characterized not by change, but by stasis.’ (Emphasis added.)32
In actual fact, there are some examples of erectus which display quite a large ECV, such as Vertesszöllos.33 To make matters even more interesting, there are human skulls in Australia, dated as modern, which exhibit clear and unambiguous erectus features. Found in Victoria (Kow Swamp), and New South Wales (Willandra Lakes, Mungo), several of these Australian aboriginal remains have fully modern human-sized brains of around 1250 cc, yet they all possess the heavy supraorbital tori, flattish receding foreheads, prognathic faces, and large jaws so typical of the earliest and the latest erectus specimens.
These skulls are dated from less than 15,000 years to around 35,000 years BP.34 Attwood and Edwards found it ‘… a conundrum’ that the Kow Swamp people with their more-erectus features lived later than the Lake Mungo people of New South Wales which were more ‘modern’ in appearance, and which date from around 35,000 BP.35 (See Figure 5.)36

Evolutionists Groves and Thorne have had a heated argument about the Kow Swamp puzzle,37 but neither seems to come up with a real answer as to how an erectus form persisted so late in time, and according to Chris Stringer, the presence of erectus forms in Australia — ‘… is especially perplexing, because it seems to contradict the global trend toward a more gracile skull’.38 This therefore contradicts the standard paradigm because very ‘modern’ human fossils are well known back to and beyond c.100,000 BP.39 The Australian Cossack skull, mandible and limb fragments which are dated at only 13,000 years or less, is another example of a sapiens/erectus of modern day. The widely distributed Australian fossils include an extremely robust individual, Willandra Lakes hominid 50, which is so ‘primitive’ that Thorne says, ‘… this skull is so robust it makes the Kow Swamp ones look gracile!’40 On the same page Flood acknowledges that all of these robust or ‘primitive’ people are definitely Homo sapiens, despite their possession of so many archaic features. (See also Lubenow.41)
How do evolutionist authorities reconcile the presence in Australia of both modern sapiens types and erectus types all within the last 35,000 years? One explanation given is that the erectus fossils are only late-surviving relicts of H. erectus, but this seems very doubtful seeing that erectus elsewhere in the world is supposed to have died out around 300 kya. That is a long time in which no change was evident except for the larger brain size of Kow Swamp and Cossack. Rhys-Jones believes that this ‘… extraordinary situation’ is due to Kow Swamp people being a - ‘… relict group of the original occupants of Australia … ’42 Thorne believes the two distinct Australian groups (‘modern’ Mungo and ‘primitive’ Kow Swamp) had separate ancestries -one from Java and the other from ancient China.43
Whatever explanation is offered, the stubborn fact remains that these erectus traits have persisted so long, which seems to conflict with the alleged global tendency to more gracile forms. In addition, Lubenow points out that there are 106 fossil individuals with erectus morphology which are dated by authorities more recently than 300 kya, and of these at least 62 date as recent as 12,000 BP, including Cossack, Kow Swamp and Solo Man from Java.44
Because of the alleged time factor, all these erectus-like individuals are obviously classified as Homo sapiens. Lubenow lists 16 erectus characteristics, and almost all, including brain size, are found in the above individuals. As he says, ‘ … ( erectus ) is truly a man for all seasons.’45 If these fossils dated from say 300 kya, they would undoubtedly be assigned to the taxon Homo erectus.
On the other hand, if creationists are right in believing that earth has only a short history (measured in millennia, rather than millions of years), then there would be no argument - all forms of Homo (except the phantom-like H. habilis), would form just a single contemporary species.
The growing creationist (and evolutionist) view is gathering strength - that H. erectus and all H. sapiens forms should be considered not as separate species but as a single human species encompassing a range of genetic and phenotypic diversity. In the creationist view there was no evolution from the apes,46,47,48 nor was there any phylogenetic, ‘ascent’ from an inferior type of human to a more advanced type.
There are a number of possible, even probable, non-evolutionary explanations for this diversity in the human species, and these are succinctly outlined in the works of Lubenow and Beasley.49,50
Also, Custance has argued convincingly that the so-called ‘primitive’ erectus and Neanderthal features are almost entirely due to the functioning of the jaw mechanism which would affect the size and shape of brow ridges, the forehead and the zygomatic arch.51 On page 183 Custance finds that the ‘primitive’ facial and skull features have nothing to do with evolution, but are due to the eating of uncooked or partially-cooked foods, especially in childhood, thus strengthening the jaw mechanism, causing it to be- come more massive in structure, and this process deforms the skull by depressing the forehead, making the brow ridges more prominent, and forces outwards the zygomatic arch, thus accentuating the cheek bones. If these people also chewed hides and skins of animals for softening, this would also have had a similar effect. This effect is increased by the tugging of flesh from the bones, and might be particularly pronounced when the diet, especially of juveniles, is lacking in bone-hardening substances such as calcium. By mid-adolescence these features then would become ‘set in concrete’ as adult characters.
Custance cites known examples, and points out that such authorities as Hooten, Howells, Hrdlicka and others were well aware of this.52 Thus, such a process, occurring in individuals, could well account for many erectus and Neanderthal features. Custance’ works should be compulsory reading for all anthropologists, whether creationist or evolutionist.
Origins
Over the years it has been generally assumed that A. afarensis gave rise to A. africanus and/or Homo habilis, which in turn evolved into H. erectus. About 300–400 kya it is then believed that archaic sapiens forms arose from an erectus population (see Figure 6).53
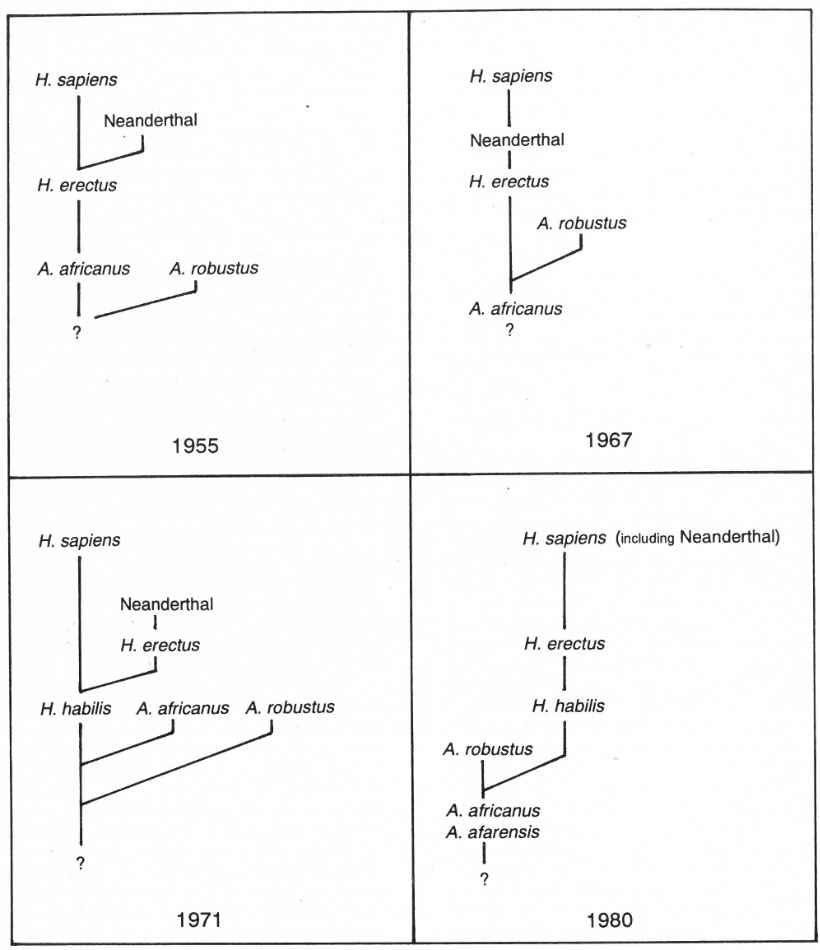
All ‘archaic’ forms display varying degrees of facial prognathism, brow ridges and brain capacity. The three best-known examples are Swanscombe (England), Steinheim (Germany) and Broken Hill (Rhodesian Man, Africa). All three also possessed flattish foreheads and ECV’ in the 1200–1300 cc range. This raises questions about the origin of Neanderthals and modern man which evolutionist authorities have so far been unable to satisfactorily solve. The matter of the origin of the archaic sapiens themselves is also unresolved. The modern Omo specimens were found close together and have identical ages; yet Omo II has noticeable erectus features, while Omo I is as modern as people of today. Wood says that if the two were found in separate locations they would have been put in different groups. This demonstrates the large range of variability in a contemporary human population.54
The fact that these forms arose well before the first Neanderthals, although many of them were allegedly more ‘progressive’ than these, is a widely-debated problem, as is the fact that the more ‘progressive’ Neanderthals with a steeper forehead appear in the evolutionary fossil record before the classic or more ‘primitive’ European Neanderthal forms, thus indicating that the former did not evolve from the latter as the morphology might otherwise suggest.
The patterns differ according to continent. The earliest archaic sapiens appear in Europe (Germany and Greece) around 600–700 kya (Petralona and Mauer, Heidelberg), around 350 kya in France (Arago 21) and Germany (Steinheim). The earliest African archaic forms are Ndutu, Tanzania (at 450 kya) and Saldanha, South Africa (at about 300 kya). The Swanscombe female archaic skull from England dates from about 300 kya. All of these bear erectus-type as well as more modern-type cranial features.
There are no unambiguous archaic sapiens in Asia but two recently-discovered skulls from China seem to have the flattened erectus-type foreheads, yet their ECV’ are apparently close to the modern human average and their faces are flatter than the usual erectus specimens. The dating is a little uncertain but the generally agreed date is around 250–300 kya. This date and the dolichocephalic configuration (narrow and oblong viewed from the top) of the skulls has led researchers to classify them both as H. erectus.55,56 ,57
Bunney reported in 1986 that a human skeleton dating from c.280 kya in China antedates an erectus skull from Zhoukoudian (the Peking Man site) near Beijing by 50 kya. The earlier skull is typical erectus in its morphology, yet has a rounded occiput and a brain case of about 1390 cc. Authorities are thus in a fix -the date says H. erectus, but the brain size is modern Homo sapiens.58 The Vertesszöllos erectus from Hungary is a very similar case.
Lubenow gives a pen-picture of the main archaic sapiens diagnostics:-
- Low, sloping forehead,
- Cranial capacity from 1100-1300c,
- Heavy supraorbital ridges,
- Facial prognathism, and
- Modern post-cranial skeleton.59
He considers that by #8216;making up#8217; this category called ‘archaic sapiens’, evolutionists wish to portray them as transitionals between erectus and Neanderthals and modern man. It is not my belief that they intend to deceive at all; this taxon could be said to represent a convenient classification for the purposes of discussion and reference, even though the overall differences are so slight. These specimens do certainly pose problems for evolutionary theory but the mix of modern and erectus characters is real enough. The main conundrum appears to be how and why so many advanced or derived characters are present so long before either Neanderthals or modern humans appear in the fossil record, and Lubenow is correct in attacking the picture by referring to the dates.60 Also, why should Neanderthal forms of less than 100 kya display so many ‘primitive’ features if it was merely a matter of straight- forward progression from erectus to archaics and Neanderthal?
Until recently there has been little sign of Neanderthals much before about 120 kya in evolutionary time, yet during the alleged period from 90,000–35,000 years BP, they undoubtedly were contemporaneous with modern men and women. The two sub-species are both found in Würm glacial deposits of the Upper Pleistocene coexisting in caves in Palestine (Skhül, Qafzeh, Tabun, Kebara), for up to 40,000 years of geological time.61 Late reports have been published which alter the picture. Richards reports from Madrid that a cave in Spain (Sierra de Atapuerca) has yielded a number of Neanderthal finds. He notes that despite these skulls being much older than any previous finds, they are indisputably Neanderthals. The finds date from well beyond 300,000 years ago- a time when Neanderthals were simply not thought to have existed - and the degree of variability in this single group of fossils, is a surprise to palaeontologists. The presence of modern features in this cave group so early, and so long before the classic type appeared only adds to the puzzle.62
Another report is by Dorozynski in Science, where it is stated that some of the skeletal features at Altamura in Italy vary enough to argue they must have belonged to at least two species, one of which led to Neanderthals, while, the other led to Homo sapiens sapiens. Yet because they all belong to one group, others claim that these variations are relatively insignificant, and the hominids all belong to one lineage.63
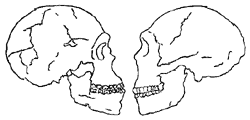
A skull from Tabun is classic Neanderthal, yet in nearby Skhül a number of skulls are intermediate between classic Neanderthals and modern humans. According to Waechter, the Skhül population may be the hybrids of Neanderthals and true modern sapiens who were already in existence.64 A gracile Qafzeh skull is quite modern with a high forehead and short braincase, whereas a Tabun skull nearby is classic Neanderthal with occipital bun; yet both had the brain capacity of fully modern humans.65 (See Figure 7.)66 While these questions pose problems for strict evolutionary progression, much of the difficulty lies in the assumption of evolution in the first place.
Lubenow’ book is the best general creationist expose of human evolution so far published, although I disagree strongly with his willingness to accept the KNM-ER 1470 skull (classified as Homo habilis) as probably human. This skull has far too many australopithecine features to be anything else than a large-brained africanus. In addition, a new reconstruction has recently been made, and an examination of the meatus angle (the pitch of the face onto the cranium) for example, shows the following: common chimpanzee 49°, A. africanus 47-53°, P. boisei 53°, and H. habilis (including KNM-ER 1470) 52–53°. The erectus skull ER 3733 shows a marked jump up to 66°, indicating that all the previous ancestors had ape-faces and no progression is seen through the australopithecines and ‘habilis.’ The angle of two habiline specimens is exactly the same as that of africanus and boisei. The craniofacial index (facial area versus cranial area) of all ‘habilines’, including 1470 falls within the australopithecine range and outside the human range including erectus.67,68
As for our putative ape-man ‘ancestor’ A. afarensis, Peter Schmid of Zurich’ Anthropological Institute recently revealed his surprise when he first examined Lucy’ skeleton (as cited by Leakey and Lewin in Origins Reconsidered). In addition to all her other ape features, it now turns out that even her rib-cage is pure ape. Schmid then turned his attention to Lucy’ entire upper body: ‘The shoulders were high, and, combined with the funnel- shaped chest would have made arm swinging improbable in the human sense… The abdomen was potbellied, and there was no waist… . In other words, Lucy and other australopithecines were bipedal, but they weren’t humans, at least in their ability to run.’69 On pages 194–196 of Origins Reconsidered we find that Aiello in London also found discrepancies: ‘No doubt about it’, she states. ‘Australopithecines are like apes, and the Homo group are like humans. Somethingmajor occurred when Homo evolved … ’ On page 195 a diagram shows clearly some of the major differences between australopithecines and humans. The reference to bipedality can be taken with a grain of salt because some previous studies have shown that it is highly likely that if Lucy did walk upright on occasions it was in the same manner as modern-day chimpanzees.70 Incidentally, computerized tomography carried out by Conroy and Vannier of the University of Washington confirms that the teeth of the Taung child (A. africanus) were developing in a distinctly ape-like manner.71
Bromage points out that the first reconstruction of ER 1470 was erroneous by giving it a flat face, but #8212; #8216;… recent studies of anatomical relationships show that in life the face must have jutted out considerably, creating an ape-like aspect, rather like the faces of Australopithecus#8217;.72 This finding is one of a number which suggest that the species Homo habilis never existed. In reality all ‘habiline’ forms display unmistakable australopithecine traits.
Lubenow’ discussion of the australopithecine, habiline, erectus and Neanderthal specimens is an excellent exposition of the creationist view that the first are extinct chimp- or gorilla-like forms, and the ‘habilines’ are nothing more than variants of australopithecines, while erectus/Neanderthal/modern man are simply varieties of a single human species.
Neanderthals — So-called
The scientific treatment of Neanderthals over most of the past century has been nothing short of scandalous. Marcellin Boule, the prominent French palaeontologist of the early 20th century, produced a very faulty reconstruction and description of a Neanderthal — an error which was to persist until after World War II.73
Lubenow cites a number of authorities who finally have acknowledged the full humanity of Neanderthal people,74 and he also claims that a Neanderthal skull has been recovered in Israel (Amud 1), which appears to date at only about 6,000 years old, although this date is controversial, as a fission-tracks test has produced an age of 28 kya.75 On pages 75–77, Lubenow cites evolutionists Klein, Geist, Angel and Wright who have all produced non-evolutionary theories which explain Neanderthal features and which are very similar to Custance’ hypothesis mentioned earlier.76 (It appears that what evolutionists see as ‘primitive’ or ape-like features are nothing more than climatic, dietary or other pathological effects on individuals of a population, allowing also for some degree of racial or genetic variation. These explanations can account for all four groups -erectus, archaic, Neanderthal and the ‘modern’ erectus forms such as Kow Swamp.)
The question of the archaic sapiens from a creationist viewpoint has been dealt with superbly by Beasley,77 who offers a cogent, reasonable and well-researched case that the archaic sapiens are our post-Flood ancestors with variable morphological features due to such influences as prolongation of skeletal maturation and greater longevity potential; environmental; and dietary/pathological pressures within a post-Deluge framework. Beasley’ paper is a land- mark in creationist research on this subject, and every Christian should possess a copy with which to question materialist-minded educators.
Trends In Secular Literature
The elevation of Neanderthal man to his rightful and proper place as a full human being was recently endorsed whole-heartedly by Rensberger. In a detailed summary, this well-known evolutionist has come to the only conclusion possible -that far from being a mixture of brute and human, Neanderthals were as human as you and I, and he is very critical of the earlier misguided and inexcusable approach which denigrated them as sub-human. Rensberger’ work should be a salutary lesson for those who are inclined to accept scientific pronouncements uncritically as being Gospel truth.78
With respect to Homo erectus, movement in evolutionary circles to upgrade his status is also gathering momentum. Peter Andrews discusses the Middle Pleistocene specimens from Java and China, and earlier Pleistocene forms ER 3733 and 3883 from Africa, and the later European and African forms such as Arago, Heidelberg and Broken Hill (Rhodesian Man).79 He claims the African specimens may represent different species or a separate lineage from Asian forms giving rise to separate populations of H. sapiens in the later Pleistocene -that is, Solo Man from Java may be directly ancestral to the controversial Kow Swamp and Cossack erectus/sapiens populations in Australia about 6,000–13,000 years ago. In Europe he speculates that the Neanderthal forms may be ancestral to modern man, and finally in Africa, a line from Ternifine through Broken Hill and Omo. (Omo 1 is virtually identical to modern man and dates from c.115–130 kya).80
If so, then H. sapiens is either polyphyletic (three independent lineages), or evolved gradually on a broad geographical front between 400–200 kya. The implication is clear: ‘… that H. sapiens and H. erectus are one and the same species which is changing gradually through time.’ (Emphasis added.)81 I agree that they are one and the same species-exhibiting variability in the early centuries after the Flood.
On page 25 Andrews also discusses the possibility that Homo sapiens evolved only once in Africa and spread into Europe twice, once giving rise to the Neanderthals, and later via Skühl and Qafzeh populations, to Homo sapiens in Europe. Yet on the same page he says: ‘ … it has been demonstrated that the European hominid sequence could be viewed as a single lineage… culminating in the classic Neanderthals of the last ice age.’ (Emphasis added).
To the informed creationist, most of this is meaningless and irrelevant. The morphological differences within all erectus specimens and between erectus, Neanderthal, and all Homo sapiens are so small that there is not the slightest reason to doubt that every form should be classified in a single human species, as we have already seen advocated.
It is not generally known to the lay-person that there are a number of modern humans who display the same erectus/Neanderthal features in addition to the Kow Swamp and Cossack examples. Taylor has published a photo of an Indian woman who clearly exhibits very large supraorbital tori.82 On the next page he gives another example of a living human who was closely examined in the Philippines in 1908. The man had massive brow ridges, a poorly defined chin, and a large lower jaw. I personally have seen and conversed with two East European immigrants where very large brow ridges and a flattened forehead were clearly visible, yet both, who incidentally were squat and heavily built, were normal individuals.
The human skull is very plastic in early childhood and I believe Custance’ theory of dietary/climatic effects may account for some erectus/Neanderthal features very well, while racial/genetic variability is also to be considered. Evolution from animal ancestors is not involved.
It used to be popular years ago to stress the distinctions between Neanderthal Mousterian tools and those of early modern man, but Wood83 admits that recent evidence indicates that these differences are not at all clear cut. Mousterian tools are found with modern skulls at Jebel Irhoud and it is therefore misleading to associate Mousterian tools solely with Neanderthal Man.84 It is also clear that not all erectus/Neanderthal specimens possessed all of these so-called primitive features, such as the Teshik- Tash 10-year-old and the Krapina ‘A’ juvenile. These were both youngsters but the 18 year-old youth Ehringsdorf ‘F’ fossil possessed both a steep forehead and a fairly high cranial vault, a moderate torus and thinner bones.85 Two Neanderthal mandibles, Ehringsdorf ‘F’ and ‘G’, an adult and a juvenile, had well -developed chins.86 In view of these by-no-means-rare cases, any evolutionary ‘progression’ simply does not fit the bill.
As Beasley points out,87 a lack of dietary vitamin D alone can account for some, but not necessarily all of the so-called ‘primitive’ features of Homo erectus and Neanderthal Man. Beasley also notes that some primitive features are still to be found in various extant racial groups.88
Wolpoff and his colleagues of the University of Michigan, because they believe the five main human races—Negroid, Caucasoids, Mongoloids, Australian aborigines and southern African bushmen—began their evolutionary divergence well before becoming anatomically modern Homo sapiens, totally reject the ‘out of Africa’ hypothesis, whereby all modern people owe their ancestry to Africa only—the Noah’ Ark theory.89
Wolpoff is supported by Alan Thorne of the Australian National University.90 According to Shipman, Wolpoff and others are now ‘… proposing nothing less than the complete abolition of Homo erectus on the grounds that the species is insufficiently distinct from Homo sapiens. All fossil specimens of Homo erectus and archaic Homo sapiens (including Neanderthals), … should be reclassified into a single species, Homo sapiens, that is, subdivided only into races.’ (Emphasis added throughout.)91
Under the Wolpoff/Thorne scheme the new definition of Homo sapiens would include all human ancestors with brain sizes from 850–2000+ cc. Of course this would totally exclude the australopithecines and the phantom ‘habilis’— position which creationists would thoroughly endorse. Wolpoff and Thorne argue (correctly) that H. habilis is too morphologically distinct from both erectus and sapiens and therefore should be excluded from the genus Homo.92 John Reader has also outlined many of the problems facing ‘habilis’, and concludes: ‘… more than twenty years of accumulating evidence and discussion have left Homo habilis more insecure than it ever was.’93 Creationists again would agree because it seems obvious that ‘habilis’ is only an australopithecine ape. Wolpoff and Thorne cannot find any consistent anatomical markers which separate erectus from sapiens. They point to the mix of sapiens and erectus features in the two recently discovered Chinese fossil skulls which virtually proves that erectus and sapiens are members of the same species and the taxon Homo erectus should be laid to rest.94
Other authorities such as Rightmire disagree, claiming that the minor distinctions which Wolpoff et al. consider as merely racial variations, are sufficient to keep separate species classifications.95 On these same pages, Shipman points out the difficulties in identifying meaningful points for measuring skull vault thickness for example. The variation, individual to individual, is considerable and this is exactly one of the points which I am attempting to make- ‘evolution’ has nothing to do with it. The differences between the various forms of archaic Homo sapiens relates, at least in part, to a combination of climatic, dietary, maturational and longevity-driven factors. In a short article in 1990, Maslen cites Dr Thorne as saying -
‘… the fossil record reveals that the features possessed by the early hominids who lived in Europe, Asia and Africa, have exactly the same sort of range as those we see in modern people.’96
I have made no attempt to enter the molecular debate and the Noah’ Ark controversy. This subject has been covered by creationist authorities.97,98 Michael Denton, a non-creationist, has already indicated the non-reliability of the molecular clock.99
Dr Carl Wieland has drawn attention to articles published in Germany, and sent to Australia in November 1992. According to these reports a well preserved erectus-type skull has been uncovered in a gravel pit near Reilingen. Although originally discovered in 1978 its significance was only realized several years later. If the dating is correct, this erectus specimen must have coexisted with modern (quasi-archaic) sapiens; the age corresponds with that of Steinheim and Swanscombe. Another article from Die Welt of September 27, 1986 refers to this ‘Homo erectus reilingensis’ and another erectus fossil from Bilzingsleben allegedly dating from about 300 kya, and there is evidence of stone tools and implements made out of elephant bone and antlers. The details of the Reilingen skull portions indicate a mix of erectus and more modern features, thus confirming the hypothesis set forth in this paper.100
Summary
In a previous article101 I demonstrated the lack of suitable primitive ancestors for the australopithecines, for ‘habilis’ and for Homo erectus. In this paper I hope to have shown that the erectus-archaic-Neanderthal–modern man ‘chain’ is non-evolutionary; that is, that all these forms are simply varieties of human beings (see Figure 8).102 The question of time is irrelevant—it makes no difference whether the time involved is millions of years or only a few thousand. The morphological distinctions are very much insufficient to warrant placing these forms in separate categories which only serves the purpose of evolution - a theory to which so many scientists are committed as dogma. Once evolution is accepted as dogma, all evidence is interpreted in a subjective, rather than objective, manner. Creationists will continue to expose the weaknesses in the theory and to encourage further research and study into the origins of man. It is not simply a matter of blind adherence to religion; the evidence is available to all who honestly wish to study and evaluate it.

The actual facts of the fossil record, that is the fossil materials themselves as against evolutionary interpretations of these materials, show indisputably that contrary to expectation the ‘earliest’ erectus skeleton (WT 15000 or the Turkana ‘Boy’) proves by its very existence that this human being was large like modern humans, and not small and ape-like. On the other hand fossil OH 62 proves that ‘habilis’, far from being Homo-like, was small and ape-like - these cases were the very opposite of what evolution theory predicted and expected.103 Even though the brain size of WT 15000 was smaller than most modern humans, it was still larger than quite a few people living today.
As for time and geology, all known facts are subject to somebody’ interpretation. Some are reasonable, others are not. It is reasonable for example to interpret an inferior rock layer as having been deposited earlier in time than the layer above, but when it comes to the question of how much earlier all estimates must necessarily be based on a number of assumptions, and this also applies to all radiometric and geological dating methods. The estimates made may be correct, partially correct, or totally wrong, and therefore almost all of geology, time and fossils are exposed to subjectivity, no matter how carefully the experts tackle their tasks. Once evolution is accepted as dogma all evidence is interpreted in that light.
Nobody can be totally certain by scientific methods alone, as to the reason why fossils are often found in certain patterns - was it by natural causes of slow deposition over long time periods as life-forms evolved, or by equally natural causes over a short time-frame such as by a global flood (and/or subsequent residual catastrophes) depositing the life-forms which were biogeographically zoned? No scientist was there to observe what happened, but others (Noah and his family) were eye-witnesses and have given us an account of the Flood.104,105
Lay-persons reading popular science books, magazines or newspaper articles will assume that the picture of human evolution presented therein is a depiction of established fact. This view is totally erroneous - the scientists have a scanty, but in places reasonable, selection of past life-forms—50 or 60 pages of a book of unknown length, and if (as usual) they are already believers in evolution, these representatives of past life will inevitably always be interpreted in the light of the theory.
Other alternatives such as presented here and in various creationist works are equally valid, but it is emphasized that neither creation nor evolution can be scientifically proven.
After careful study of hundreds of scientific descriptions, and photographs of scores of fossil humans, it is clear to me that all shades of intergrading exist between ‘ancient’ erectus and modern humans, but the chronological patterns of appearance, even using the evolutionists’ own dating methods, do not match the predictions of the theory. In view of the clear-cut and unmistakeable morphological gap between apes and humans, I believe that human fossil study provides strong circumstantial evidence in favour of the theistic view of origins as outlined in the early chapters of the book of Genesis, as against the current view that random or chance genetic accidents were responsible.
The late A. W. (Bill) Mehlert had a Diploma in Theology and lived in Brisbane, Australia. He was a keen student of Flood geology and the fossil record, including the supposed fossil evidence for human evolution, and wrote a number of important articles on these topics in the Creation Research Society Quarterly and Journal of Creation.
Acknowledgment
The drawings on which the figures are based were prepared by my son Gary Mehlert.
References and Notes
- Whitcomb, J. and Morris, H., 1961. The Genesis Flood, Baker Book House, Grand Rapids, Michigan. Return to text.
- Mehlert, A W., 1992. A review of the present status of some alleged early hominids. J. Creation (formerly CEN Technical J.) 6(1):10–41. Return to text.
- Gish, D. T., 1985. Evolution: The Challenge of the Fossil Record, Creation-Life Publishers, El Cajon, California, pp. 140-180. Return to text.
- Lubenow, M.,1992. Bones of Contention, Baker Book House, Grand Rapids, Michigan, pp. 50–57, 157–168, 172–179. Return to text.
- Beasley, G. J., 1990. Pre-Flood giantism: a key to the interpretation of fossil hominids and hominoids. J. Creation (formerly CEN Technical J.) 4:5–55. Return to text.
- Lubenow, Ref. 4, p. 127. Return to text.
- Cberfas, J. and Gribbin, J., 1981. Descent of man or ascent of ape? New Scientist, 91(1269):594–595. Return to text.
- Gish, Ref. 3, pp. 180-203. Return to text.
- Bowden, M., 1977. Ape-Man: Fact or Fallacy? Sovereign Publications, Bromley, Kent, pp. 78–148. Return to text.
- Johnson, W., 1982. The Crumbling Theory of Evolution, Queensland Binding Service, Brisbane, pp 40–45. Return to text.
- Lubenow, Ref. 4, p. 135. Return to text.
- Lubenow, Ref. 4, p. 91. Return to text.
- Fezer, K. D., 1993. Creation’ incredible witness: Duane T. Gish, Ph.D. Creation/Evolution 33:5–21 (p. 12). Return to text.
- Lubenow, Ref. 4, p. 86. Return to text.
- Brace, C. L. and Montagu, A., 1977. Human Evolution, Second Edition, Chicago University Press, pp. 204-205. Return to text.
- Leakey, R. and Walker, A., 1985. Homo erectus unearthed. National Geographic 168(5):624–629. Return to text.
- Leakey, R. and Lewin, R., 1992. Origins Reconsidered, Abacus Books, London, pp. 58–64. Return to text.
- Leakey and Walker, Ref. 16, pp. 624–629. Return to text.
- Jue, D. S., 1990. Cranial capacity and endocranial casts. J. Creation (formerly CEN Technical J.) 4:56–65. Return to text.
- Broderick, A H.,1971. Man and His Ancestors, Hutchinson and Co., London, p. 84. Return to text.
- Lubenow, Ref. 4, pp. 123, 128. Return to text.
- Beasley, G. J., 1992. Pre-Flood giantism: a key to the interpretation of fossil hominids and hominoids. Unpublished manuscript (original Version), p. 71. Return to text.
- Beasley, Ref. 22, p. 71. Return to text.
- Lubenow, Ref. 4, p. 138. Return to text.
- Jue, Ref. 19, p. 57. Return to text.
- Lubenow, Ref. 4, p. 138. Return to text.
- Molnar, S., 1975. Races, Types and Ethnic Groups, Prentice-Hall, Englewood Cliffs, New Jersey, p. 57. Return to text.
- Pellegrino, C. R., 1985. Time-Gate, Hurtling Backward Through Time, TAB Books, Blue Ridge Summit, Pennsylvania, p. 120. Return to text.
- Beasley, G. I., 1992. A possible creationist explanation of archaic fossil human remains. J. Creation (formerly CEN Technical J.) 6(2):138–167. Return to text.
- Lubenow, Ref. 4, pp. 78–85. Return to text.
- Lubenow, Ref. 4, pp. 29–30. Return to text.
- Pellegrino, Ref. 28, p. 124. Return to text.
- Campbell, B. G., 1988. Humankind Emerging, Fifth Edition, Time-Life Books, Harper ColIim, New York. pp. 287–288. Return to text.
- Attwood, A., and Edwards, K., 1987. Footprints of early man. Time Australia 2(13):28-41 (supplementary map). Return to text.
- Attwood and Edwards, Ref. 34, p. 37. Return to text.
- Wolpoff, M. and Thorne, A., 1991. The Case against Eve. New Scientist 130(1774):35. Return to text.
- Groves, C. and Thorne, A., 1991. Brawl about Eve. The Australian Magazine, February 16–17, p. 28. Return to text.
- Stringer, C., 1990. The emergence of modern humans. Scientific American 263(6):74. Return to text.
- Wood, B., 1976. The Evolution of Early Man, Cassell Australia, Sydney, pp. 98-100. Return to text.
- Flood, J., 1990. The Riches of Ancient Australia, University of Queensland Press, Brisbane, p. 20. Return to text.
- Lubenow, Ref. 4, p. 133. Return to text.
- Attwood and Edwards, Ref. 34, pp. 37–39. Return to text.
- Attwood and Edwards, Ref. 34, pp. 37-39. Return to text.
- Lubenow, Ref. 4, pp. 131–132. Return to text.
- Lubenow, Ref. 4, pp. 132-133. Return to text.
- Mehlert, Ref. 2, pp. 33-38. Return to text.
- Lubenow, Ref. 4, pp. 134–143. Return to text.
- Beasley, Ref. 5, pp. 31-55. Return to text.
- Lubenow, Ref. 4, pp. 149–156. Return to text.
- Beasley, Ref. 5, pp. 5-55. Return to text.
- Custance A., 1975. Genesis and early man. The Doorway Papers, Volume II. Zondervan, Grand Rapids, Michigan, pp. 183–184, 208–211. Return to text.
- Custance, Ref. 51, p. 184. Return to text.
- Campbell, Ref. 33, pp. 200–201. Return to text.
- Wood, Ref. 39, p.99. Return to text.
- Gibbons, A., 1992. An about-face for modern human origim. Science 256:1521. Return to text.
- Shipman, P., 1993. On the origin of races. New Scientist 137(1856):36. Return to text.
- Tianyuan, L., and Etler, D.A., 1992. New Middle Pleistocene hominid crania from Yunxian in China. Nature 357:404–407. Return to text.
- Bunney, S., 1986. Chinese fossil could alter the course of evolution in China. New Scientist 111(1520):25. Return to text.
- Lubenow, Ref. 4, p. 80. Return to text.
- Lubenow, Ref. 4, pp. 78-85. Return to text.
- Waechter, J., 1976. Man Before History, Elsevier International Publishing Co., Oxford, p. 64. Return to text.
- Richards, M., 1993. Caveman shows his face. The Courier Mail, Brisbane, November 8, p. 9. Return to text.
- Dorozynski, A., 1993. Possible neandertal ancestor found. Science 262:991. Return to text.
- Waechter, Ref. 61, p. 63. Return to text.
- Bar-Yosef, O. and Vandermeersch, B., 1992. Modern humans in the Levant. Scientific American 268(4):68. Return to text.
- Bar-Yosef and Vandermeersch, Ref. 65, p. 68. Return to text.
- Bromage, T., 1992. Faces from the past. New Scientist 133(1803):32–35. Return to text.
- Hummer, C. C., 1977. A plea for caution about skull1470. Creation Research Society Quarterly 14(3):171. Return to text.
- Leakey and Lewin, Ref. 17, pp. 193–194. Return to text.
- Cherfas, J.,1983. Trees have made man upright. New Scientist 97:172–178. Return to text.
- Bromage, Ref. 67, p.33. Return to text.
- Bromage, Ref. 67, p. 35. Return to text.
- Campbell, Ref. 33, pp. 378–379. Return to text.
- Lubenow, Ref. 4, pp. 61–64, 76–77. Return to text.
- Lubenow, Ref. 4, p. 73–74. Return to text.
- Custance, Ref. 51, pp. 183–184, 208–211. Return to text.
- Beasley, Ref. 29, pp. 138–167. Return to text.
- Rensberger, B., 1981. Facing the past. Science ’81 2(9):41–50. Return to text.
- Andrews, P., 1984. The descent of man. New Scientist 102:24–25. Return to text.
- Wood, Ref. 39, p. 99. Return to text.
- Andrews, Ref. 79, pp. 24–25. Return to text.
- Taylor, I.,1987. In the Minds of Men, IFE Publishing Co., Toronto, p. 214. Return to text.
- Wood, Ref. 39, p. 108. Return to text.
- 84. Wood, Ref. 39, p. 108. Return to text.
- Beasley, Ref. 29, pp. 144–145. Return to text.
- Beasley, Ref. 29, p. 145. Return to text.
- Beasley, Ref. 29, p. 143. Return to text.
- Beasley, Ref. 29, p. 163. Return to text.
- Shipman, Ref. 56, p. 34. Return to text.
- Wolpoff and Thome, Ref. 36, pp. 33–37. Return to text.
- Shipman, Ref. 56, p. 34. Return to text.
- Shipman, Ref. 56, p. 36. Return to text.
- Reader, J., 1990. Missing Links, Penguin Books, London, p. 189. Return to text.
- Shipman, Ref. 56, p. 36. Return to text.
- Shipman, Ref. 56, pp. 36-37. Return to text.
- Maslen, G., 1990. Man or ape? The Sunday Mail, Brisbane, May 20, p.11. Return to text.
- Wieland, C., 1992. No bones about Eve. Creation Ex Nihilo 13(4):20-23. Return to text.
- Beasley, G. J., 1992. Is the African ‘Eve’ misconceived? J. Creation (formerly CEN Technical J.) 6(1):42–48. Return to text.
- Denton, M., 1985. Evolution: A Theory in Crisis, Adler and Adler, Bethesda, Maryland, pp. 233–307. Return to text.
- Wieland, C., 1993. Personal communication. Return to text.
- Mehlert, Ref. 2, pp. 10–41. Return to text.
- Campbell, Ref. 33, p. 388. Return to text.
- Reader, Ref. 93, pp. 188–189. Return to text.
- Woodmorappe, J.,1983. A diluviological treatise on the stratigraphic separation of fossils. Creation Research Society Quarterly 20(3):133–185. Return to text.
- Whitcomb and Morris, Ref. 1. Return to text.
Readers’ comments
Comments are automatically closed 14 days after publication.