

Journal of Creation 25(1):69–76, April 2011
Browse our latest digital issue Subscribe
The origin of the Carboniferous coal measures—part 2: The logic of lycopod root structure
New fossil and field evidence relating to the structure of the root system of lycopods, the dominant vegetation of Upper Carboniferous strata, are presented and critically examined. Neither the elastic and partially hollow nature of the lycopod root structure, their inferred geometry throughout early ontological development, nor other evidence support the prevailing paradigm that the coal measures formed in a terrestrial swamp environment. Rather, they favour the floating forest or silvomarine hypothesis of Kuntze regarding the formation of Paleozoic coal layers.
Are lycopods structured for water immersion?
Part 1 of this paper traced how researchers have traditionally thought the Paleozoic coal deposits of the northern hemisphere formed in swamps.1 It described how in recent decades there has arisen renewed interest in an alternate view—i.e. that they were the result of beached floating ocean forests. The originators of the floating forest paradigm were biologists, arguing strongly from paleontological evidence that the dominant Paleozoic fern-tree plants were structured for a water environment. An in-depth and independent investigation would be needed to counter their conclusions, but such an investigation might serve to strengthen and extend their proposals if they were correct.
In this article the author reproduces some salient points brought up by the floating forest advocates while adding her own independent observational checks and calculations. Mathematical modeling of lycopods is initiated here by looking at their root systems. Some startling and significant finds have resulted from this analytical investigation.
What do Paleozoic fern-tree roots say about coal formation?
The first thing to note about the lycopod root system is that it is found fossilized in an amazing variety of distinct rock types. Stigmaria are found in clay, claystone, shale, sandstone (including greywacke), limestone, and even coal.2 If Stigmaria were fossilized in situ as the dominant paradigm assumes, then they were lithified in a wider range of ‘soils’ than any known plant can tolerate. Also, stigmarian rootlets were designed to be shed.3 Hence, either Stigmaria were the roots of a rather unusual and extraordinary plant, or the assumption that their fossilization always took place in situ needs revision. One hypothesis that accomplishes this revision is that Stigmaria were part of a floating forest, an ancient quaking bog, in a marine environment that was rapidly buried. If the biological principle that “form indicates function” has any real meaning, Stigmaria should have morphology suited to an aquatic environment that should still be evident in their fossils.
New evidence for the vacuous nature of Paleozoic lycopod roots
|

|
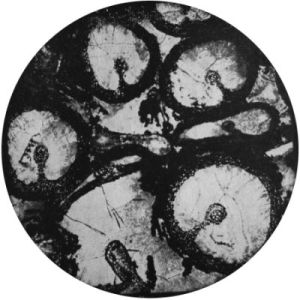
If lycopod roots were hollow, this would be a strong argument they were suited for a watery environment. Indeed, it has long been known that their appendices or rootlets were hollow. Past examination of excellently preserved rootlets in coal balls has established this fact (figure 1).4 Furthermore, vintage scientific reports have noted that Stigmaria themselves were either hollow or had interiors with contents that were easily invaded. One particularly good fossil sample collected in the past had layers in its interior that even had fern fossils between them (figure 2).5
As part of this investigation, the author has collected and measured samples of Stigmaria. All the samples discussed in this paper were collected at locations within a three-kilometer radius of a point centered just west of the commercial centre of Portersville, Pennsylvania, primarily from a sandstone layer immediately below the Middle Kittanning Coal. The fossil sites were found in the following manner. Satellite photographs were used to geomorphologically locate rapidly eroding areas on the edge of steep gullies (i.e. buried runoff channels were sought as possible Stigmaria collecting localities). The steepness of the gullies was a consequence of the stream base level being influenced by what is interpreted as a glacier-caused stream reversal. The rapid erosion was due to a perched water table, a consequence of a thin clay layer at the base of the undercoal sandstone. All predicted sites yielded Stigmaria. However, those Stigmaria that had been compressed into the thickness of a double ‘bark’ were excluded from this in-depth investigation. The existence of such samples, however, goes a long way toward proving the conjecture that they were either hollow or at least easily flattened.
Some of the samples examined strengthened this impression of an easily invaded interior. Each of these shows the broken extension of the rootlet attached to its central core or ‘stele’ swept back along and parallel to the stele’s axis as if the Stigmaria had been intrusively filled by sediment originating from a source closer to its trunk. (Evidence of prominent micaceous layers on or in the interiors was found in 23.6% of the samples6 collected and analyzed.7) In order to further test whether stigmarian interiors were hollow, measurements were taken of the first 30 logical salient characteristics of stigmarian fossils suggested by examination of the fossil samples collected.6 Among the measurements tabulated were those of their cross-sectional shape (table 1).
Table 1. Shapes of collected and measured Stigmaria from Portersville, Pennsylvania, USA.
| Cross-sectional shape | % of samples | % with steles | % with lower side covered with stigmata |
|---|---|---|---|
| Circular or Elliptic | 25.8 | 83 | 62 |
| ‘I-beam’-shaped | 3.4 | 33 | 67 |
| ‘U’-shaped | 24.7 | 45 | 51 |
| Semicircular | 38.2 | 44 | 1.2 |
| Triangular | 2.2 | 50 | 0 |
| Rectangular | 5.6 | 40 | 20 |

Many (25.8%) of the measured Stigmaria were circular or elliptic in cross-section. Moreover, 3.4% had an ‘I-beam’ shape, as if the sides had been crushed from an originally circularly-shaped object. An additional 38.2% were somewhat semicircular, as if they had been sheared (which, judging from the absence of stigmata on the base of these Stigmaria, appears to have been the case).8
The crushed ‘I-beam’ shape would be expected if Stigmaria were reinforced by spoked rootlets radiating from a stele and pinned at the root’s outer edge, and then subjected to crushing pressures. If a Stigmaria, which was originally circular in cross-section, were subjected to uniform crushing pressure from above (figure 3), it would tend to first take on an elliptic cross-sectional shape. However, this would make some of the rootlets attached along the center of the flattened top and bottom limp, while others at the ends, where the curvature was greatest, would transmit extreme tension to the sides. It would then either collapse inwards if the sides were pliable enough, or the roots or rootlets could rupture, both of these cases being observed.9 This perceived observational agreement is mathematical: all the I-beam shaped Stigmaria follow the analytical considerations contained in the preceding reference when they are being crushed, lending credence to the conjecture.10

The Stigmaria collected in Pennsylvania appear to have been easily bent, crushed, or wrinkled (figures 4, 5a, and 5b. Figure 5b is an enlargement of a part of figure 5a). Wrinkles paralleling the stigmarian axis showed in 8.4% of the measured samples. Conversely, when the longitudinal axes of the Stigmaria are bent, they exhibit compression folds along their inner radius while appearing stiffened by wrinkle-smoothing tension along their outer radii of curvature. This reflects the roots’ lateral stiffness, despite their transverse susceptibility to being crushed. Stigmaria under bending tension consistently show their steles as having moved off centre toward the outer edges in response to this bending. This bespeaks of a probable mobility of the stele, one that may indicate that the inner pith around it is very pliable or non-existent.

|

|

More telling are the locations of steles on collected samples relative to the cross-section of the Stigmaria.11 These steles should be clustered toward the centre of the cross sectional area of Stigmaria if the root were solid, instead they are bunched toward the bottom (figure 6).12 One unusual stigmarian fossil sample found in shale illustrates many previous points made above and partially settles whether or not the stigmarian cores were hollow. This meter-long mudstone or sandstone sample (missing the middle third of its length) was found in situ in the centre of the carbonaceous Clarion Shale of western Pennsylvania next to many fossil ferns (in all probability where the Scrubgrass Coal would have been if it were locally present). It constitutes the very tip of the stigmarian root. It is circular or elliptic in cross-section but progresses to a crushed ‘I-beam’ shape while its stele progressively sags as it moves towards the tree-fern trunk. More importantly, this one Stigmaria sample apparently underwent a different taphonomic (probably electrochemical) history from all other collected or viewed samples. Besides apparently lacking any trace of external rootlets, this well-preserved sample had at least 30% of its core filled with what appears to be a fossilized spongy material, possibly reproducing tracheids.13 Whatever fossilized biological material there was appears to be clustered around the central stele. So it appears, from an examination of this sample, that there could be several possibilities for Stigmaria:
- the spongy material was agglomerated toward the tip and decreased toward the trunk;
- it was pervasive near the trunk and decreased toward the tip;
- it filled the whole inside of the Stigmaria (except for the obvious places like rootlet extensions and the space occupied by the stele);
- or it filled a given percentage of the interior (30% or more).

The stele itself appears to be subject to collapse, although not nearly as readily as the Stigmaria proper, judging from the samples collected in western Pennsylvania. A progressively collapsing stele has been found on a Stigmaria from Plainville, Massachusetts. Other researchers have found indentations in the exterior shape of Stigmaria apparently mirroring delayed stele collapse (figure 7)—a phenomenon observed by the author on a specimen she did not collect, one not from her collecting region. Such a phenomenon probably indicates a different stratigraphic or taphonomic history from that undergone by all the specimens collected by her. Summarizing, it appears that every aspect of lycopod roots indicate they were both light in weight and elastically pliable beyond the normal limits generally ascribed to the roots of terrestrial tree-sized plants. This is especially true of its rootlets or appendices.
The structure and fossilization of Paleozoic lycopod roots
Stigmarian roots have long been recognized as being fossilized with their rootlets radiating out perpendicularly from them (figures: 8, 9, and 10).14 This layout is seen as an aquatic adaptation, not a terrestrial one. There appears to be no logical reason for the existence of perpendicularly radiating rootlets or appendages of lycopod roots if they belong to terrestrial plants. There are especially no reasons for them to double back upon themselves or wrap around the Stigmaria. Yet this is seen frequently in samples collected, leading to serious doubts about them being terrestrial in habitat (figure 11).15

|

|

|

|
Root systems vary widely in their form and show adaptations to markedly different life cycles. They also tend to fall into categories that can be related to their seed form. For example, most monocotyledonous plants have fibrous root systems. Those species of monocotyledons that have bulbs, corms, or rhizomes additionally have mainly adventitious roots, as do grasses where these supplement their seminal roots:
“The root systems of dicotyledonous plants, on the other hand, are usually based on a taproot and its branches. The taproot often does not persist, however, and then the framework of the root system consists of several of the lateral branches.”16
Stigmaria, the root of lycopods, does not fit into either of these categories. It has lateral roots but never a taproot. It most resembles, by analogy of form, the root systems of pond plants like the bladderwort (Utricularia vulgaris).17 The very large rootlets of Stigmaria not only stick out straight from the root radially on all sides, but they are also very long. They are so long, in fact, that the top ones would be sticking up in the air unless the lycopod they are associated with germinated from a deeply buried site or was later completely buried by sediment. Both of these possibilities are very unlikely: the lycopods’ peculiar ontogeny would seem to argue convincingly against them.
The size of this quandary can be illustrated by a few examples. The mature Stigmaria in figure 12 are all a meter or less below the germinating point of the lycopod, whereas rootlets on some similarly mature lycopods are as much as 2.0 meters long.18 There is a possible reconciliation to this apparent difficulty. If the rootlets of the lycopods were similar to the roots of mangrove trees, they would be expected to be air-breathing, possessing special tiny pores (or lenticels) and large air spaces (or aerenchyma). No explicit mention or any evidence of the existence of lenticels on the rootlets of Stigmaria have been found.

However, many cases of silica infilling the rootlets were found, as if they were transporting this mineral in later taphonomic decay. These cases are especially associated with rootlets that show bending near the root—a generally atypical occurrence. From this and other evidence, the author surmises that lycopod rootlets were stiffened by silica (as are some parts of contemporary marsh plants). This would explain their assumed high specific gravity (based on the assumption that they were neutrally buoyant). It would also protect them from rot and salinity. Their taphonomic decay rate is inferred to be the determinant of the various fossil forms these rootlets take. They usually appear as rigid and uniform cylinders radiating straight out from the root, but sometimes they appear flattened and expanded except near the root connection, or as silica cylinders with coal macerals in the middle replicating former pliability even near the root connection.
One final point should be noted concerning the rootlets. Consider the two samples with an abrupt angular flexure of many straight-sided radiating rootlets which were collected in the eastern Pennsylvanian anthracite region. These samples of flexure, not faulting, suggest some sort of mass readjustment of a semisolid block within a more fluid medium. (If it were an artefact of terrestrial erosion, the flexed rootlets would not be as straight and parallel as they are.) The majority of the author’s fieldwork was done in bituminous regions. Not a single case of slumped rootlets was found in this region. The author’s collection of Stigmaria in anthracite regions has been at least an order of magnitude less than that in bituminous ones. However, it is precisely in these regions of greater metamorphism that the two samples of slumped rootlets were found. This may be coincidence, or it may be related to some other phenomenon or combination of phenomena (e.g. more intense folding, differences in dewatering or diagenesis).

An unusual ontogeny
The lycopods had a much different development than other tree-sized seed plants. (We know this because of the painstaking work of D.A. Eggert, who serially sectioned many individual lycopod parts from coal balls, resulting in a detailed hypothetical reconstruction of their ontogeny.19) They started life as a closed megaspore, a sort of boat seed case, something one would expect if they germinated in an aquatic medium.20 Then they went on to form a young plant with a rounded stump-like, full-girth appearance (figure 13). This developmental pattern markedly differs from that of other tree-like plants, the growth of which is primarily upward with girth expansion occurring over time by the accumulation of secondary xylem (the woody element of such trees). The issue of whether or not this lycopod development reflects a design for an aquatic environment will be covered in a future article.
If a lycopod were to start its development in marine surroundings, one would suspect that it would be so designed as to be both partially sticking out of the water and also able to right itself if overturned in the juvenile stage. The known stigmarian structure, including its interior voids or spongy areas, ensures the first condition under any reasonable circumstances, by way of its presumed density.21 Let us now consider its ability to right itself.
If one imagines an ideal immature lycopod turned upside down and totally immersed in ocean water, it becomes obvious such a circumstance would be highly unstable. Because the lycopod was assumed to be originally buoyant, its four starting roots would stick out of the water. But now that they were exposed in air, they would gravitationally present a highly unstable (or metastable) situation. A root, then a second one, would fall to the water surface with the slightest perturbation.

Assuming the ontogeny of Eggert,22 there would be at least two critical design criteria that logically would have to be met if the lycopod were to right itself in order to survive. If it were overturned and were lying with two roots on the surface of the water and two sticking up in the air, then the angle between the plane of those roots on the surface of the water and the plane of those sticking up in the air definitely would have to be less than 90°. Otherwise the overturned position of the baby lycopod would be irreversible (figure 14).23 On the other hand, if it were laying somewhat sideways while two of its roots were on the water surface plane with the other two submerged—not an unreasonable situation if it were naturally righting itself—then the angle between the two planes containing the roots should be near 90° if the lycopod were to continue to right itself by way of its density distribution (including voids and spongy areas causing positive buoyancy). These two considerations need to be checked against available evidence. Together they restrict the interplane angle between pairs of roots to be close to, but less than, 90°.
It has been asserted that the rootlets shed themselves uniformly in a non-traditional manner seemingly unrelated to water availability, more like the shedding of leaves due to maturing plant growth. Shedding of rootlets is well known among plants.24 However, the rootlets referred to as being shed here are dwarfed by stigmarian appendices. Furthermore, the shedding being referred to is done by areas, the plant abandoning non-productive rootlets generally from mineral-depleted areas. This is unlikely to be the case for water emplacement (e.g. in a swamp), where mineral mobility in the fluid would tend to be equalized over larger areas. Note that the shedding of rootlets on an immature lycopod would be advantageous in its ability to right itself in a fluid environment if the rootlets shed were the expected middle ones—the ones not on or near its top or root-tip-growing surfaces. If rootlets were likely to be injured drastically enough to allow water invasion, and if they were close to neutrally buoyant to begin with, then it would make perfect sense from the stability point of view for water-immersed plants to shed them.
One lycopod root system documented in the literature (figure 15a) is close enough to the stage right before it starts its upward trunk growth that its geometry at that point could be reasonably estimated by visual backtracking of its apparent growth. When this is done and the angle between any two planes, each containing two of its primary roots, is calculated, a value of 84° is obtained.25 This lycopod root is quite typical of the Pennsylvania strata. It is of a Stigmaria ficoides Brongniart: it represents the overwhelming majority type of lycopod roots from the Pennsylvanian. On the other hand, the most unusual lycopod-like root base from the Pennsylvanian known to the author is Stigmariopsis Grand’Eury (shown from a hypothesized reproduction of it in figure 15b26). Calculating the angle between the two planes containing pairs of its immature roots gives a value of 86°. These two examples are considered as bracketing what would be expected from such root systems. Note that both of these bounding cases clearly meet the criteria necessary for lycopods to be able to right themselves if they ended upside-down in seawater. Also observe the stigmarian-like root structure shown in figure 16 of a Mesozoic lycopsid from Vladivostok, Russia.27 Its form suggests adaptation for a watery environment and further strengthens and extends the argument that many ancient plants were created for such an environment.

|

|

|
Conclusions
There is much fossil evidence suggesting the roots of Upper Carboniferous fern trees were quite elastic. In addition, there is a plethora of observations suggesting they contained vacuous and fragile organic matter, perhaps even being partially hollow. Their unusual ontogeny and radiating rootlets also suggest immersion in a watery environment. These factors provide strong evidence for the silvomarine hypothesis of Kuntze concerning the nature of Upper Carboniferous floating forests. They herald the need for uniformitarians to revisit and revise their long-age, vegetative swamp environment explanations for the formation of Paleozoic coal layers.
Acknowledgement
Thanks to Barry Lee Woolley, who assisted with some of the mathematical calculations.
References and notes
- Woolley, J.F., The origin of the Carboniferous coal measures—part 1: Lessons from history, J. Creation 24(3):76–81, 2010. Return to text.
- Kuntze, O., Geogenetische Beitrage, Gressner and Schramm, Leipzig, Germany, pp. 54–55, 1895; Lesquereux, L., Description of the Coal Flora of the Carboniferous Formation in Pennsylvania and throughout the United States, volume 1, Board of Commissioners for the Second Geological Survey, Harrisburg, Pennsylvania, pp. 510–513, 1880. Kuntze records researchers having found Stigmaria in slate clay, granite, gneiss, mica slate or micaceous schist and other silicate stones, greywacke, and various kinds of limestone, including a Devonian one. The author has also found samples in each of the lithified ‘soils’ mentioned in the text except limestone. Return to text.
- Scheven, J., The Carboniferous floating forest—an extinct pre-Flood ecosystem, CEN Technical Journal (Journal of Creation) 10(1):70–81, 1996 (see especially figure 4 on page 75 showing a fossil rootlet in the process of being shed). Also see Frankenberg, J.M., Abscission of lateral appendages of Stigmaria, American Journal of Botany, 1 January 1968. Return to text.
- Gürich-Gothan, from Hirmer, M., Handbuch der Paleobotanik, Band 1: Thallophyta, Bryophyta, Pteridophyta, R. Olderbourg, Berlin, p. 294, 1927. The fossil Stigmaria collected by the author in western Pennsylvania often had distinct silica rootlets emerging from them, sometimes encasing coal macerals (the different constituents of coal, analogous to minerals in rocks) that gave the impression of being in the process of migrating out from the root. The locations of macerals on the cross sections of Stigmaria will be schematically illustrated and commented upon in a forthcoming article. Return to text.
- Brown, R., Quart. Jour. Geol. Soc. Lon. 4:49, 1847. The samples were from Joggins, Nova Scotia, Canada. Return to text.
- Barry Lee Woolley, Daniel A. Woolley, C. Joy Woolley, Joshua A. Woolley, and Peter A. Woolley assisted with the collection of some of the stigmarian fossil samples. Only the first 106 of them (the average length of which was 22.6 cm) were examined in detail in order to try to avoid skewing results by over-sampling one locality or one fern-tree. Return to text.
- The muscovite mica appears not to be metamorphic in the least, both from texture examination and fluvial structural association. Weaver, C.E., Beck, K.C., and Pollard Jr., C.O., Clay Water Diagenesis during Burial: How Mud becomes Gneiss, Geological Society of America, Boulder, Colorado, p. 21, 1971. Return to text.
- These high percentages of seemingly crushed Stigmaria seem to be characteristic of the author’s collecting locality in western Pennsylvania but not necessarily of all other collecting sites. The stigmata on stigmarian fossils are sometimes partially or wholly absent due to weathering, which is reflected in the values in the last three rows in the last column of table 1. Return to text.
- If the Stigmaria were to be completely crushed, the original angle theta (measured in a Cartesian coordinate system centered on the stele and referenced to the Stigmaria’s major horizontal axis in cross-section) where a rootlet would not have been subjected to any limpness or extensive forces would be just under 33º. More precisely, it would be the complement of one radian. This angle was easily obtained by finding the distance along the Stigmaria’s circumference, starting from where the vertical axis cuts its upper surface, that is equal to its radius. Return to text.
- If the number of rootlets around the periphery of a Stigmaria that was being crushed were ‘n’, and if they were numbered starting on the
extreme right and proceeding counterclockwise, then the place of no tension, the place that lies nearest the inward bend in an ‘I-beam’-shaped Stigmaria, will be
the numbered rootlet as given by the following table: Nearest numbered peripheral stigmarian rootlet to the start of the inward bend.
If the number of peripheral rootlets along the whole circumference is ‘n’ Then the rootlet nearest the start of the bend (first quadrant only) is 0=n Zeroth one 1<n<5 First one 6<n<16 Second one 17<n<27 Third one 28<n<39 Fourth one
Note that the author has also found samples of fossil Stigmaria with strips of the bark bent back upon themselves. Return to text. - In a 27 January 2006, personal conversation with Howard R. Shanks of Ames, Iowa (who has for years collected Pennsylvanian fossils at Pella, Iowa, and sold them at the annual Tucson, Arizona, commercial fossil shows), he asserted that the steles, when they exist, in all of the Stigmaria samples he has ever collected have always been at the bottom of the cross-section. He also asserted that none of the Stigmaria he has collected showed the ‘I-beam’ cross sectional shape: they have always been round or elliptic. All his fossils were in sandstone. The fern tree spacings he has witnessed he estimated to be fifteen feet (or up to twenty feet) (4.6 up to 6.1 meters) apart at this Iowa location. Return to text.
- In figure 6, which shows the stele locations on the cross-sections of Stigmaria collected by the author, there may be some inadvertent or systematic clustering errors for off-major-axis reporting of stele locations. This is because contorted, creased, and elliptic stigmarian cross-sections meant angular measurements were only estimates in a number of cases. Nevertheless, the clustering of steles towards the bottom of Stigmaria fossil samples is a universal observation not at all peculiar to the western Pennsylvanian location from which the measured ones were taken. Return to text.
- The electrochemical functions performed by rootlets in attracting minerals makes the absence of rootlets coupled with the presence of a normally missing spongy interior less surprising. Return to text.
- The radial spread of rootlets or appendages perpendicularly outward from Stigmaria appears to be an uncontested fact. The author has plenty of evidence that this should be the case. However, there are significant and wide variations from this normal pattern. These variations include, for example, rootlets lying along the stigmarian axis. Return to text.
- Not many fossil Stigmaria samples that were collected had extant rootlets attached to them: most radially spreading rootlets were left with the matrix from which the Stigmaria were extracted. Return to text.
- Head, G.C., Shedding of roots; in Kozlowski, T.T. (Ed.), Shedding of Plant Parts, Academic Press, New York, p. 239, 1973. Return to text.
- The perpendicularly outward-radiating rootlets attached to a stele (generally fairly straight and sometimes suffering from distinct angular junction bends of their straight-sided rays) as opposed to exceedingly pliable rootlets wrapped along a stele or undulating outward from it seem contradictory but are observational facts. This may be related to the fact that the pliable rootlets are more likely to be associated with flat sheets of lycopod bark, crushed or creased or broken Stigmaria, and increased distance from the stele. It is guessed that the taphonomy undergone by the rootlet is the basic cause of the seemingly huge increase in pliability. Furthermore, it is pure speculation that stigmarian stiffness is related to high silica content, which would be rather unusual for terrestrial plant roots or rootlets (as opposed to other parts of these plants). Return to text.
- Kuntze, ref. 2, p. 48. Return to text.
- Eggert, D.A., The ontogeny of Carboniferous arborescent lycopsida, Palaeontographica B 108:43–92, 1961. Return to text.
- Warwick, P.D. (Ed.), Coal Systems Analysis, Geological Society of America, Boulder, CO, p. 46, 2005. The lycopod dense bark-like tissue, called the periderm, was waterproof and decay resistant. More about lycopsids, Devonian Times, www.devoniantimes.org/who/pages/lycopsid.html, accessed 22 November 2010. Return to text.
- Samples of its original woody structure, which could be bent or burned in a fire just like ordinary wood, have been recovered. Hence it appears unreasonable to assume that its density would lie outside that of the contemporary range of woody plants. Dawson, J.W., Quart. Jour. Geol. Lon. 2:132–134, 1846. Return to text.
- Other assumptions were (1) lycopod wood density was close to that of contemporary wood (i.e. less than that of water); (2) root voids distributed so as to have the leaves on top of the plant out of the water with the roots in it (i.e. air voids lying between the root tips and the genesis point); (3) rootlets that are neutrally buoyant; and, of course (4) stigmata space separation between rootlets and leaves. Some of the last of these assumptions help, but are not essential to, the argument. Return to text.
- The analysis was started with the Stigmaria precisely upside down in the water, finding that a spread >90° would prejudice and doom the root from falling on its side and beginning to aright itself, respectively. Angles <<90° would doom the plant from moving off its side position to begin its final righting phase. Note that in the case of two roots being in the air, using natural righting requires the extra weight of the non-submerged rootlets for its successful completion under reasonable immature lycopod weight distribution assumptions. Return to text.
- Head, ref. 16, pp. 237–293. Return to text.
- The angles between the planes for both examples are calculated as follows: the three components of vectors along each of the four roots are measured off of the appropriate views given in figure 15a. These four vectors are then made into unit vectors by dividing by their lengths. Unit normal vectors to the two planes each containing two roots are subsequently formed from vector cross-products of each of the two sets of paired unit vectors along the roots. Next, the vector dot product is used to obtain the angle between the planes through an inverse cosine calculation. Note that a reference in the book containing both of these examples claims the angles between the planes typically lie between 70° and 80°, despite the photographs and what the author has calculated. See Hirmer, ref. 4, p. 285. Return to text.
- Hirmer, ref. 4, p. 297. In order to represent only its early development, its length is considered terminated on each of its four roots at the midpoint of where the second set of ‘taproots’ emerges. Return to text.
- Hirmer, ref. 4, p. 326. Return to text.
- Gürich-Gothan of 1923; from: Hirmer, ref. 4. Return to text.
- Brown, ref. 5, p. 49, fig. 7. Return to text.
- Cleal, C.J and Thomas, B.A., Plant Fossils: The History of Land Vegetation, The Boydell Press, Woodbridge, Suffox, UK, 1999. Return to text.
- Williamson, W.C., A Monograph on the Morphology and Histology of Stigmarian ficoides, Palaeontological Society of London, 1887; as reproduced in: Hirmer, ref. 4, p. 287. Return to text.
- Williamson, ref. 31, p. 326. Return to text.


Readers’ comments
Comments are automatically closed 14 days after publication.