

Journal of Creation 29(3):62–70, December 2015
Browse our latest digital issue Subscribe
What life is
The materialistic view of life as a natural phenomenon has been deeply contradicted by research into its molecular mechanics. In this article I briefly survey progress towards an explanatory theory of life based on molecular mechanics. The main proposition is that sustainable life is irreducibly cellular, and cell structure is irreducibly and continuously hierarchical and cyclic throughout its history in 4-dimensional space-time. One proof of this proposition lies in the cell wall. A primordial cell is irreducibly dependent upon its protective cell wall, which is irreducibly hierarchically and cyclically structured, and it is irreducibly dependent upon the cell contents for its topological continuity throughout its history in space-time. The cell contents, by the same argument, must have the same properties. Primordial life must be astonishingly complex: autotrophic, using Brownian machines to exploit thermal noise in quantum-engineering top-down-designed self-templated self-assembling substructures, while intelligently managing information exchange to ensure rational decision making in maintaining homeotic balance in the face of continuously conflicting demands from internal and external environments. Irreducible continuity of cell structure and function throughout its history in space-time defies naturalistic explanation. Only Genesis-style fiat creation can explain it.
Life is astonishing in its structure, function, and capabilities. Most professional scientists believe its origin lies in some chance combinations of aqueous organic chemistry, so they expect it to exist wherever there is liquid water in the universe.1 Organic chemist Addy Pross, in his 2012 book What is Life? How chemistry becomes biology, gave an apparently sophisticated explanation by reverse engineering contemporary life back through imagined evolutionary time.2 But this is self-delusion—it simply affirms what the author assumed (chemical evolution) and assumes what must be explained (cell structure, function, and capability). Nobel Prize winning biochemist Christian de Duve was a rare exception in acknowledging the pervasive obstacles in this worldview.3
In contrast, astrobiologist and philosopher of science Carol Cleland has observed that we need to know what life is before we can hope to explain its origin, and to do that we need “a general theory of living systems”.4 Harvard University’s Nobel prize-winning origin-of-life researcher Jack Szostak has concluded that cells are the essential prerequisite:
“… the question we’re looking at is what do we need to do to make these chemicals get together and work like a cell?”5
Like others, Szostak began at the bottom and worked up towards increasing complexity and functionality, but without success (also like others). He then decided to study the transition to cellularity by building an artificial cell.6 However, Williams7 has demonstrated that the primary structure required by a primordial cell is a strong cell wall to protect it from the destructive thermal energy of free water (called the ‘molecular heat storm’). This has profound consequences, and in this article I review some of them to illustrate what life is, and thereby contribute to the development of a general theory of living systems.
The cell wall
A brief overview of different kinds of life demonstrates that all cells need protection from the destructive power of free water. Prokaryote cells (figure 1A) are protected by a strong fibrous or paracrystalline capsule. Eukaryote unicellular amoeba walls (figure 1B) have a majority composition (~75%) of strong molecules (proteins and phosphoglycans) interwoven through their cytoplasmic membrane to strengthen it while maintaining flexibility so the amoeba can crawl in complex ways to find and ingest food particles and avoid predators.8
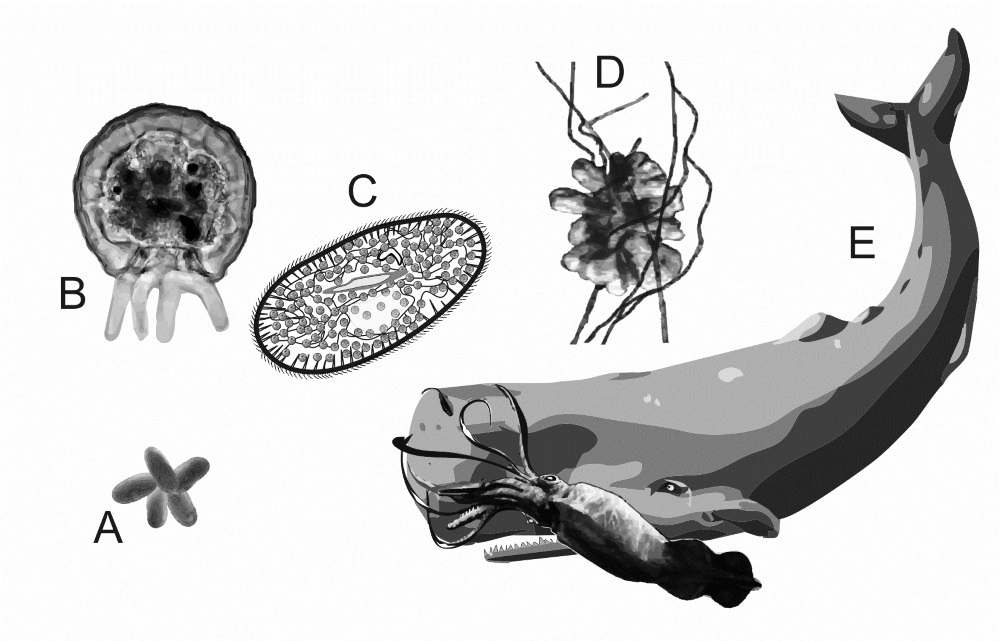
Unicellular paramecia (figure 1C) have a stiff, flexible, skin-like pellicle that protects the cytoplasmic membrane, which then overlays a polygonal network of fibres which anchor their bodycovering cilia.9 Multi-cellular plants (figure 1D), together with algae and fungi, have strong fibrous cell walls made from a variety of polysaccharides, including cellulose and chitin.10 Multi-cellular animals (figure 1E) house their cells within a flexible, fibrous, extra-cellular matrix, which is thickened on the outside to produce a leathery skin. Unprotected animal cells in blood are kept safe through the blood serum being concentrated enough to neutralize the cell’s osmotic pressure gradient. If blood is diluted with too much water, the cells burst.
Prokaryotes must be primordial in naturalistic scenarios because they are much simpler than eukaryotes. The capsule which protects the prokaryote could be compared to something like a leather football (figure 2A). The ball has an impervious rubber bladder that holds the contents (compressed air) while an outer leather casing protects it from rupture when kicked. The prokaryote cell has an inner cytoplasmic membrane that guards the cell contents, and an outer capsule that protects it from rupture when exposed to the molecular heat storm (figure 2B). Prokaryotes have several variations in their wall structure and three are illustrated. Gram-positive bacteria have an inner phospholipid bilayer membrane and multiple strong layers of peptidoglycan fibres on the outside (figure 2C). Gram-negative bacteria have a phospholipid bilayer membrane outside as well as inside, with a strong peptidoglycan fibrous layer between them (figure 2D). Archaea have a phospholipid bilayer membrane on the inside and a paracrystalline pseudopeptoglycan strong layer on the outside (figure 2E).
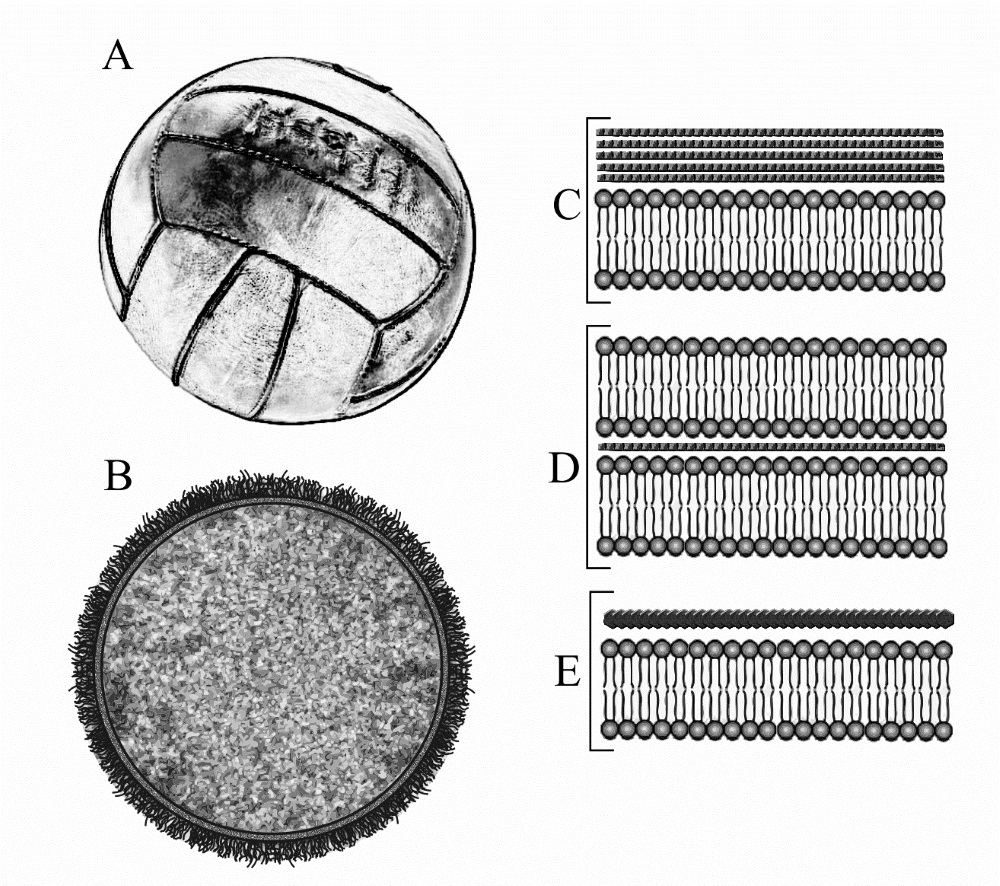
No prokaryote capsule lacks a strengthening layer. This proves that lipid bilayer membranes alone are of no use to cells in the natural environment, yet they are the universal starting point in origin-of-life experiments. It’s a fanciful delusion.
Cell wall construction
One of many stumbling blocks encountered by Szostak’s team in constructing artificial cells is that the capsule holding the cell contents must expand as the cell grows, and it must divide when the cell divides. Synthetic capsules, like the plastic ones containing oral medications, are fixed in size—they do not grow or divide. So how does the prokaryote capsule do it? The same way that most of the other cell components do it—by ‘self-templated self-assembly’.
To understand this remarkable mechanism (and its relevance to the origin of life) we need to look at a brief description of how, and why, humans use it:
“Fabrication of [molecular] architectures from topdown technology involve[s] precise growth techniques like molecular beam epitaxy, chemical vapor deposition and also involve[s] patterning techniques such as photolithography, particle beam lithography, scanning probe lithography, and nanoimprint lithography. While the above mentioned processes are laborious, time-consuming, and costly, the ‘bottom-up’ technology based on [the] self-assembly approach is the simplest, cost effective technique. Self-assembly is one of the most important ‘molecular engineering’ strategies used in fabricating complex functional structures, from micro to the molecular levels, utilising the advantage of self-interaction of molecules. Molecular self-assembly is a strategy for nanofabrication that involves designer molecules and supramolecular entities so that molecules naturally aggregate into specific desired structures. This method reduces many difficult steps in nanofabrication … . Moreover, molecular self-assembly tends to produce structures that are relatively defect-free and self-healing, because the target structures are selective with thermodynamically stable assembly between the possible configurations [emphases added].”11
The gulf lying between a prokaryote capsule and an artificial capsule is that the prokaryote version is dynamic, undergoing continual turnover. As one end of a wall polymer self-assembles new material, the other end is enzymatically dismantled and recycled. When growth is required, an up-regulation of monomer supply at the growing end is sufficient to ensure the wall elongates in the right direction to accommodate the expanding cell contents. The pre-existing wall polymers act as templates on which the new material self-assembles. This has profound consequences for cell origin because it means that new cell walls can grow only from pre-existing cell walls. Self-templated self-assembly cannot arise out of a ‘blank slate’ origin, it can only continue from the wall of a pre-existing cell.12 None of this is possible with artificial capsules.
As you can see from the quotation above, self-templated self-assembly is not a naturally occurring phenomenon. It requires “designer molecules and supramolecular entities” which can self-assemble at the right place because “the target structures are selective with thermodynamically stable assembly”. That means the molecular machinery which accomplishes self-templated self-assembly is specially designed so that the statistical mechanics of physics and chemistry ensure that the single correct configuration is automatically chosen from the myriad wrong ones. Self-templated self-assembly is an ideal method of ‘bottom up’ construction, but it only works when the whole system is intelligently designed to function that way.
The cell contents
The constraints that the above facts place upon the cell contents also have profound consequences. The cell contents must be assembled in their entirety before the cell wall is sealed, otherwise the molecular heat storm would destroy it. Cell machinery is made of large polymers and they are prevented by the selective cytoplasmic membrane from getting into the cell after the wall is sealed off. Having everything ready at the beginning requires ‘top-down’ design. So it is not just the cell wall that requires top-down design but the entire cell contents also. Human engineers don’t build bridges by randomly assembling bits of steel on river banks. If they did, either the resulting structure would fail under load, or it would be washed away in the next flood, or it would corrode into uselessness before anything functional emerged from its haphazard disarray. Instead, engineers first consider the challenges to be overcome, then plan a set of structures that will meet those needs. Then structures are made and assembled according to the plan. This is top-down design and construction.13
With the cell contents safely inside the protective wall capsule, we can now begin to think about them in their entirety. Such an enterprise goes far beyond a journal article but some idea can be obtained from EcoCyc, the E. coli modelling project. The ‘Metabolic Cell Overview’ option provides a zoomable diagram of the bacterial cell with metabolic pathway annotations.14 I will consider just some of its general principles.
One of the first points that Nobel Prize winning physicist Erwin Schrödinger identified in his 1944 book What is Life? was that the laws of physics and inorganic chemistry are based on the statistical mechanics [random interactions] of large numbers of atoms and molecules, and they have no power to explain the extraordinary behaviour of living organisms. Of special interest to Schrödinger (given the technological limitations of his time) was the curious behaviour of chromosomes during reproduction. Only organic chemists had made any progress by uncovering the remarkable structures of biochemical molecules and some of the metabolic cycles that are unique to life.15 Schrodinger thus identified that it was the unique structures of biomolecules that provided clues to the distinctive functions of living organisms.
In 1968 Michael Polanyi elaborated on this principle by noting that life’s machines displayed irreducible hierarchical structure and this was the key to understanding their functions.16 However, Schrödinger ingeniously calculated that cellular machinery must function at a scale where the thermal energy of its atoms and molecules (the ‘molecular heat storm’) constantly interferes. This changes the game entirely from what Polanyi envisaged. To solve this problem, Schrödinger proposed that strong molecular bonds must hold large molecules together, and that these large molecules must have the ability to make quantum-like jumps between different stable states. He then formulated a definition of life as “that which avoids rapid decay [through thermal degradation of its structures] into the inert state of equilibrium [where function ceases]”.17
With modern atomic force microscopes (AFMs) we can now see Schrödinger’s predictions regarding thermal energy fulfilled throughout the cell.18 Furthermore, the most important functional ‘strong molecules’—proteins and DNA—do indeed undergo quantum-like jumps between stable states. Proteins can undergo allosteric conformational changes, and DNA can change its fourbase coding sequences. But even more spectacularly, crucial events in the cell are now known to involve real quantum state transitions.19 Implications for the origin of life are staggering.
Human excursions into the quantum world usually require enormous effort and cost. Particle accelerators have to generate extremely high energies and temperatures to access the subatomic world. Quantum superconductivity requires extremely low temperatures and ultra-pure materials. But protein machines in a cell can tap into the molecular heat storm and use its incessant chaos to access quantum transitions without changing the temperature a single degree! Humans can engineer quantum phenomena at room temperature too, but it requires the highest standards of ingenuity, design, construction, and operation of equipment.20
Life’s dependence upon quantum engineering is all pervasive. To avoid thermal degradation, all cells need continuing supplies of energy and nutrients, and they must extract these from the environment. Autotrophs—cells that can manufacture food from sunlight, inorganic chemicals, or electrons—are the only kind of cells that could have been primordial. Heterotrophs—cells that rely on organic food from outside (e.g. primordial soup)—would soon have exhausted their local supply and died out. Earth’s biosphere is maintained by a vast network of autotrophs, providing food for heterotrophs, then recyclers turn it all back into raw materials to be used over and over again. Any sustainable biosphere must do likewise.
One well-studied autotrophic prokaryote, Rhodospirillum rubrum, a purple proteobacterium commonly found in mud, gives some idea of what is required. It can grow aerobically or anaerobically, in dark or light, extracting energy via cellular respiration, fermentation, photosynthesis, or photoautotrophic growth. A supercomputer simulation of its chromatophore harvesting light reveals a marvel of quantum-inclusive engineering.21 During photosynthesis light energy is converted into chemical food energy via a ‘special pair’ of chlorophyll molecules embedded slightly off-centre to one another in a surrounding protein matrix. Their mutually interacting but ‘out of tune’ vibrations allow them to amplify quantum interference effects that ‘tune out’ wasteful energy transfer routes and ‘tune in’ only the most efficient ones. The resulting energy efficiency is almost 100%, far exceeding the theoretical limit set by the laws of thermodynamics.22
Proteins are fundamental to all life, and their structure and function both depend upon quantum engineering. Proteins are made up of long chains of amino acids strung together via distinctive ‘peptide bonds’. These bonds require special enzymes (also made of proteins) to make them in the necessary presence of (otherwise destructive) water,23 and also to break them (for repair and recycling). They use “precision engineered” equipment to access unstable quantum-transition states and achieve what would otherwise be impossible.24 Furthermore, all cells depend heavily upon electron transfers, and it appears that proteins are a major component of cellular electronics. They can access quantum critical states and behave as semi-conductors, which are the centrepiece of our electronic devices.25
Systems biology
Many biologists now realize that life only makes sense as a whole. ‘Systems biology’ takes reductionist research results and puts everything together to build a big picture of how organisms work as a whole. The crucial difference between a living process (e.g. protein production from DNA) and a living system is that the living system has a cyclic structure. Proteins require DNA to define their amino acid sequence, but DNA also requires protein to yield up its coded information that defines the amino acid sequence. The protein production stage is entirely useless without the information management stage. Both are required concurrently for either stage to be functional.
But over the top of this particular cycle there must also be a genomic control process that regulates gene activity by determining which genes are switched on at any one time and which are switched off. The gene regulatory process must also be cyclic because everything a cell does is repeated continuously over various timescales, culminating in the reproductive cycle. However, feeding into every level of this complex cycle-of-cycles there needs to be energy, nutrient, and manufacturing supply chains; removal of waste products; and component building, maintenance, repair, and recycling routines. All of these subsystems must also be cyclic because the output of any one routine is always the input to another one somewhere else, and all the pieces need to be broken down and recycled back into the system so that waste does not build up to toxic levels. Nothing works alone. Everything must work together. And it all needs to be up and running in full functionality before it is sealed inside the primordial cell wall. It takes about 1 nanosecond for free water molecules to cross the space inside a prokaryote cell, which means the contents must be assembled on at least a picosecond timescale.

The complexity of life’s cyclic processes (figure 3A) is so great that we easily lose sight of their essential simplicity. A cycle is topologically equivalent to a circle (figure 3B). In a circle there is no beginning and no end. Each point on the circle is continuous with the ones either side of it. Each point is as necessary as every other point.
Life is not just a collection of special molecules, it is a hierarchically integrated, robust, and self-regulating system. Some first steps in capturing its complexity can be found in The Handbook of Systems Biology: Concepts and Insights.26 The most important point the book makes is that life functions only as a system.27 Michael Savageau, Distinguished Professor of molecular biology at UC Davis, has recast Theodore Dobzhansky’s famous statement about evolution into this new light: “Nothing in biology makes sense except in the light of systems.”28
A modern-day smart phone provides a useful analogy for the holistic nature of systems biology. When we press the ON button, amazing things happen because thousands of lines of coded information inside the phone spring into action. The nearest wireless telephone tower is activated and messages from all around the world flow into the palm of your hand. The smart-phone provides a galaxy of electronic technology packed small enough to hold in one hand, but big enough to enable watching of TV and movies. Now compare that with a cultured stem cell from a Tasmanian blue gum tree. Apply a drop of cytokinin and a young gum tree begins to grow, but it has no roots. Transfer the tiny tree to a new medium that has auxin in it and the roots begin to grow. After further appropriate care it can be planted out into the wild and grow up to become one of the largest hardwood trees in the world. Now take a fertilized egg from a zebrafish and apply BMP4 to one end and Nodal to the other end. Just like pressing the ON button on the smart phone, the embryo turns into a fish (it does take a bit longer). One button to activate a smart phone and two ‘buttons’ to activate a gum tree and a fish. Everything needed to produce all the action is packaged into the hand-held phone, and likewise into the tiny single cells that produced the living organisms. Life comes as a complete package in just one, whole, tiny cell.29
Cells are intelligent
A crucial property of living systems identified by the founder of General Systems Theory,30 Austrian biologist Ludwig von Bertalanffy, is that they maintain themselves very far from physical equilibrium in open systems that cannot survive apart from their environments. This property has profound consequences. Cells must always remain closed to the environment to protect their contents from hazards, but simultaneously must remain open to their environment for material and information exchange. To maintain a homeotic balance in the face of these continually changing and conflicting challenges, cells must make rational decisions which minimise the risks and maximise the benefits of every transaction.
In 1983 microbiologist James Shapiro published an article entitled “Variation as a genetic engineering process”.31 Over the next three decades he developed this idea into the concept of ‘natural genetic engineering (NGE)’—the notion that cells (not genomes) are in control of life, and that bacteria in particular are intelligent agents in charge of their own metabolism, social history, and hereditary potential.32 It was most fully explained in his 2011 book Evolution: A view from the 21st century.33 It drew heavy criticism from neo-Darwinists, but Shapiro is confident their theory has been soundly refuted.34 In a clarification of what “natural genetic engineering does and does not mean” he said this:
“NGE describes a toolbox of cell processes capable of generating a virtually endless set of DNA sequence structures in a way that can be compared to erector sets, LEGOs, carpentry, architecture or computer programming. NGE operations are not random. Each biochemical process has a set of predictable outcomes and may produce characteristic DNA sequence structures. The cases with precisely determined outcomes are rare and utilized for recurring operations such as generating proper DNA copies for distribution to daughter cells [emphasis in original].”35
In other words bacteria can engineer their genomes in different ways for different purposes. They can copy them with 100% accuracy when required, and they can dice and slice and mix them with foreign DNA of any and every kind. Shapiro concludes that “bacteria are the most successful cell biologists on the planet”.36 He has now been joined by a range of other biologists in different fields making similar claims that cells are intelligent agents.37,38,39
The ‘attention schema’ theory of animal consciousness illustrates how intelligence works. It consists of a three-tiered hierarchy.40 At the base is a network of sensory inputs. In the middle is an information processing system. And at the top is a holistic response mechanism. Holistic response does not occur after every sensation but is triggered only under certain circumstances. Those circumstances have to be mediated (but not necessarily determined) by the information processing system. In humans, conscious awareness is a whole-of-brain experience that focuses attention upon just some sensory inputs but not others.41 The subconscious information processing system determines which stimuli are brought to conscious awareness, then the consciousness determines what the holistic response will be. Bacteria do not have human-like self-awareness, but they certainly do have bacterial self-awareness. Just one individual bacterium can do everything described here, which means it must be doing so for its own sake. Its behaviour is not merely some emergent property of group dynamics.
Bacteria also have a three-tiered hierarchy of sensory input, information processing, and holistic response.42 They have thousands of receptor molecules in their outer walls and can sense dozens of different kinds of stimuli, including chemical, mechanical, biological, heat, light, and vibration sources. Memory and a sense of the flow of time are essential components. Cells maintain short-term memory regarding increasing and decreasing trends in both internal and external conditions,43 and long-term memory in regard to diurnal cycles and cell history, including periods of asexual and sexual reproduction and dormancy. Cells can count numbers of molecules and measure concentrations of chemicals, and they can do calculations, including ‘greater than,’ ‘less than’, and ‘equal to’ comparisons. They can also do Boolean algebra in ‘if … then’ logical circuits using operators, including AND, OR, and NOT. Then they integrate all this data in ways that produce rational decision making (e.g. swimming towards food and away from toxins, deciding when to cooperate and when to compete with neighbours).
Cell decision making is much more than built-in algorithmic reaction. When faced with multiple conflicting signals and internal demands, multiple possible threats, multiple courses of possible action, and multiple possible outcomes, we can observe individual prokaryotes making decisions that humans would agree are rational in the circumstances. Shapiro admits that we do not yet know how they do it but: “[It] almost certainly … [is] more than a strictly mechanical process. … [it] will certainly involve cybernetics. … [and the mechanism may be] more than strictly material … .”35
Intelligences communicate. For example, to remain healthy every cell in a multi-cellular organism must cooperate with its neighbours. This is achieved via a multiplicity of signalling and receptor pathways—the cells must share and respond to neighbourhood information. If cells break the cooperation rule it results in diseases like cancer and autoimmune disorders. Cooperation is just as necessary in prokaryotes when one cell divides and produces a dense colony of offspring,44 and when one species cooperates with other species to form the complex biofilms that are active agents in ecosystems45 and pathogenesis,46 and in the consortiums which achieve biochemical reactions not possible in component species.47 Bacteria can communicate with other members of the same species, with members of different species, and with higher organisms, including plants, animals and fungi. A rapidly increasing list of completely different kinds of organisms is now known to communicate with one another. Vastly different organisms routinely live intimately together or inside one another, sharing common resources, including information via cell signalling. Most plants actively engage with microbes, including fungi, in their root zones to complement (or even replace in some orchids and cycads) the normal functions of their root systems. The human body is host to a vast and growing catalogue of species. Cloning experiments show that cells of quite different species can be grown peaceably together in one body. Healthy human organs can grow in the bodies of pigs, sheep, rabbits, and mice.48 Human genes can operate in the bodies of plants, yeast, fruit flies, and bacteria. The language of life—not just reading the genetic code, but perceiving and acting in accordance with living systems—appears to be universal.
Prokaryotes thrive in every moist environment on earth, demonstrating beyond doubt that they are supremely good at being aware of, responding to, and communicating with their surroundings in a rational manner.
Genetic entropy
Cellular machines use the laws of physics, but they must also obey them. The fact that they live among and use thermal noise has earned them the name ‘Brownian machines’.49 Amazingly, their most energy-efficient point to operate is where their frequency of vibration matches the noise.50 The ‘dance of life’ is choreographed on the edge of chaos,19 and this may allow them greater flexibility than if they operated in a calmer environment.51 Not surprisingly, damage is frequent and much of the cell’s machinery is devoted to maintenance and repair (especially DNA) and efficient breakdown and recycling of worn-out parts. Lifetimes of RNA and proteins in bacteria are measured in just minutes.52 Mutation is more a result of physics than biology.53 A detailed study of the 1918 influenza virus genome showed that the mutations which caused its extinction (twice) were “overwhelmingly the product of thermodynamics”.54 The main biological contribution is that natural selection can remove the rare lethally deleterious mutations, but not the common sublethally deleterious ones. Since the latter are the vast majority they accumulate and multiply across generations, degrading fitness to the point of extinction.55,56 Genetic entropy confounds Darwinism, sets a short timescale for life on earth,57 and demands the highest possible primordial engineering standards.58
Topology of the cell

Topology is a branch of mathematics that deals with continuity.59 Metabolic cycles in a cell are continuous entities and topologically equivalent to 2-dimensional circles, but since they always cycle over time they are more like three-dimensional helices. Similarly, the prokaryotic capsule is topologically equivalent to a 3-dimensional sphere, like the football in figure 2, but it is also dynamic in spacetime so it is more like a 4-dimensional hypersphere. Because the capsule must always protect the cell contents from the molecular heat storm, but is itself always dependent upon the cell contents for its maintenance, growth and division, it must have topological continuity throughout its history in spacetime. Since the cell contents are always dependent upon the capsule for protection, they too must have topological continuity throughout their history in spacetime. Figure 4 contains some topologically continuous objects that illustrate these concepts.
The simplest continuous object is a 1-dimensional line (figure 4A). Increasing in dimensions and complexity are a plane (4B) and a cylinder (4C). Figure 4D, a Mobius strip, symbolizes how the cell wall on the outside is necessarily continuous with the inside metabolism. Figure 4E, a Klein bottle, makes the same point with a container. Figure 4F is a simple backwards projection of a sphere to symbolize that the cell is continuous back through time. Figure 4G, a partial view of a Hopf Fibration,60 is a mathematically accurate mapping of a 4-dimensional hypersphere into 3 dimensions, and it thus gives a more accurate representation of the cell’s continuity throughout its history in spacetime.
Conclusions
Life is irreducibly cellular, and cells function only as whole systems, continuously protected from the environment inside a strong cell wall. The necessity for the cell wall to be sealed from the beginning against the external violence of the molecular heat storm has spectacular consequences. The cell contents must be assembled in complete functionality inside the cell wall before the cell wall is sealed, and the cell, as a unit, must remain continuously functional throughout time (apart from special cases of dormancy) for life to avoid thermal decay. Yet cells also need to remain continuously open to their environment for material and information exchange. Managing a homeotic balance in the face of these challenges requires intelligent sensation of both internal and external conditions together with rational decision making. Primordial cells must also be autotrophic. Achieving these things concurrently requires the highest standards of molecular and quantum-mechanical engineering skill at ambient temperatures and picosecond timescales. No naturalistic scenario has ever come even near to meeting such criteria. Yet, despite these high standards of technological excellence, genomes are decaying on timescales of just thousands of years.
This very brief characterization of life can be conveniently summarized in the topological concept of continuity in all essential features throughout its history in 4-dimensional space-time. Continuity is an absolute criterion—it must be present from the beginning or the cell is destroyed by the molecular heat storm. Continuity cannot be added at some later stage to a ‘porous bag’ of ‘sloppy molecules’ in a step-wise Darwinian process of ‘bottom-up’ assembly over many generations. The materialistic view of life as a natural phenomenon is indefensible. Only Genesis-style fiat creation can explain it.
References and notes
- Irwin, L.N., Méndez, A., Fairén, A.G. and Schulze-Makuch, D., Assessing the Possibility of Biological Complexity on Other Worlds, with an Estimate of the Occurrence of Complex Life in the Milky Way Galaxy, Challenges 5:159–174, 2014 | doi:10.3390/challe5010159. Return to text.
- Pross, A., What is Life? How chemistry becomes biology, Oxford University Press, 2012. Return to text.
- de Duve, C., Singularities: Landmarks on the Pathways of Life, Cambridge University Press, 2005. Return to text.
- Mullen, L., Defining Life, Astrobiology Magazine, 19 June 2002, astrobio.net. Return to text.
- Making Life from Scratch: Biochemist Jack Szostak’s Search for the First Cell (2012), origins.harvard.edu, 17 June 2014. Return to text.
- Blain, J.C. and Szostak, J.W., Progress Toward Synthetic Cells, Annual Review Biochemistry 83:11.1–11.26, 2014. Return to text.
- Williams, A.R., What Life Isn’t, J. Creation 29(1):108–115, 2015. Return to text.
- Korn, E.D. and Wright P.L., Macromolecular Composition of an Amoeba Plasma Membrane, J. Biological Chemistry 248(2):439–447, 1973. Return to text.
- Sedar, A.W. and Porter, K.R., The fine structure of cortical components of Paramecium multimicronuleatum, J. Biophysical and Biochemical Cytology 1(6):583–604, 1955. Return to text.
- Domozych, D.S. et al., The cell walls of green algae: a journey through evolution and diversity, Frontiers in Plant Science 3:Article 82, 2012. Return to text.
- Pradeesh, K. et al., Naturally Self-Assembled Nanosystems and Their Templated Structures for Photonic Applications, J. Nanoparticles vol. 2013, Article ID 531871, 2013 | dx.doi.org/10.1155/2013/531871. Return to text.
- Oliferenko, S. et al., Positioning cytokinesis, Genes & Development 23:660–674, 2009. Return to text.
- Burgess, S., Hallmarks of Design: Evidence of purposeful design and beauty in nature, 2nd edn, Day One Publications, Leominster, pp. 30–31, 2004. Return to text.
- Cellular Overview of Escherichia coli K-12 substr. MG1655 (EcoCyc), ecocyc.org, 20 August 2015. Return to text.
- Schrödinger, E., What is Life?: The Physical Aspect of the Living Cell, Cambridge University Press, ch.2,1944. Return to text.
- Williams, A.R., Life’s Irreducible Structure—Part I: Autopoiesis, J. Creation 21(2):109–115, 2007. Return to text.
- Schrödinger, E., What is Life?: The Physical Aspect of the Living Cell, Cambridge University Press, 1944, ch. 6. Return to text.
- Hoffmann, P.M., Life’s Ratchet: How Molecular Machines Extract Order from Chaos, Basic Books, New York, 2012, Kindle Edition. Return to text.
- Al-Khalili, J. and McFadden, J., Life on the Edge: The coming age of quantum biology, Bantam Press, London, 2014. Return to text.
- Yirka, B., Demonstration of room temperature spin transport in germanium, 25 May 2015, phys.org/news, accessed May 2015. Return to text.
- Chandler, D., Strümpfer, J., Sener, M., Scheuring, S. and Schulten, K., Light Harvesting by Lamellar Chromatophores in Rhodospirillum photometricum, Biophysical J. 106(11):2503–2510, 2014. Return to text.
- Al-Khalili and McFadden, ref. 19, p. 127. Return to text.
- Sarfati, J., Origin of life: the polymerization problem, J. Creation 12(3):281–284, 1998; creation.com/polymer. Return to text.
- Al-Khalili and McFadden, ref. 19, ch. 3. Return to text.
- Hewitt, J., Quantum Criticality in life’s proteins, phys.org/news, 13 April 2015. Return to text.
- Walhout, A.J.M. et al. (Eds.), Handbook of Systems Biology: Concepts and Insights, Academic Press, London, 2013. Return to text.
- Carvunis, A-R. et al., Interactome Networks; in: Walhout, A.J.M. et al. (Eds.), Handbook of Systems Biology: Concepts and Insights, Academic Press, London, ch. 3, 2013. Return to text.
- Savageau, M., Phenotypes and design principles in system design space, hh.15, p. 288; in: Walhout, A.J.M. et al. (Eds.), Handbook of Systems Biology: Concepts and Insights, Academic Press, London, 2013. Return to text.
- Williams, A.R., Heredity is foundationally cellular, not genetic, and the history of life is discrete, not continuous, J. Creation 28(3):73–79, 2014. Return to text.
- von Bertalanffy, L., General System Theory: Foundations, Development, Applications, George Braziller, New York, 1968, revised edition 1976. Return to text.
- Shapiro, J.A., Variation as a genetic engineering process. In Evolution from Molecules to Men (D.S. Bendall, Ed.), Cambridge University Press, pp. 253–270, 1983. Return to text.
- Shapiro, J.A., Bacteria are small but not stupid: cognition, natural genetic engineering and socio-bacteriology, Studies in the History and Philosophy of Biology & Biomedical Sciences 38:807–819, 2007. Return to text.
- Shapiro, J.A., Evolution: A view from the 21 st century, FT Press Science, Upper Saddle River, NJ, 2011. Return to text.
- Natural genetic engineering, en.wikipedia.org, accessed August 2015. Return to text.
- Shapiro, J.A., What Natural Genetic Engineering Does and Does Not Mean. Posted: 28 February 2013, huffingtonpost.com. Return to text.
- Shapiro, J.A., Inter-Kingdom Horizontal DNA Transfer in All Directions: Infectious Bacteria Evolve by Acquiring Protein Domains From EukaryoticHosts, updated 6 January 2013, huffingtonpost.com. Return to text.
- Trewavas, A., Plant Behaviour and Intelligence, Oxford University Press, 2014. Return to text.
- Mancuso, S. and Viola, A., Brilliant Green: The surprising history and scienceof plant intelligence, Island Press, Washington D.C., 2015. Return to text.
- Karban, R., Plant Sensing and Communication, University of Chicago Press, 2015. Return to text.
- Graziano, M., How consciousness works, Aeon Magazine, 21 August 2013, aeon.com. Return to text.
- Moran, M., Network theory sheds new light on origins of consciousness, medicalxpress.com, 11 March 2015. Return to text.
- Bray, D., Wetware: A computer in every living cell, Yale University Press, New Haven, 2009. Return to text.
- Lambert, G, and Kussell, E., Memory and Fitness Optimization of Bacteria under Fluctuating Environments, PLoS Genet ics 10(9): e1004556 | doi:10.1371/journal.pgen.1004556, 2014. Return to text.
- Shapiro, J.A., Multicellularity is the rule, not the exception: Lessons from E. coli colonies; in: Shapiro, J.A. and Dworkin, M. (Eds.), Bacteria as Multicellular Organisms, Oxford University Press, pp. 14–49, 1997. Return to text.
- Gautam, C.K. et al., Review Article: An insight into biofilm ecology and its applied aspects, International J. Pharmacy and Pharmaceutical Sciences 5(4):69–73, 2013. Return to text.
- Kostakioti, M. et al., Bacterial Biofilms: Development, Dispersal, and Therapeutic Strategies in the Dawn of the Postantibiotic Era, Cold Spring Harbour Perspectives in Medicine 3:a010306, 2013. Return to text.
- Shapiro, J.A., Evolution: A View from the 21st Century, Pearson Education, Kindle Edition, 2011, p. 101. Return to text.
- A good review of chimeric research and its ethical implications is at: Human/Non-Human Chimeras, Stanford Encyclopedia of Philosophy, plato.stanford.edu. Return to text.
- Guohui Li and Qiang Cui, Analysis of functional motions in Brownian molecular machines with an efficient block normal mode approach: Myosin-II and Ca21-ATPase, Biophysical J. 86:743–763, 2004. Return to text.
- Schneider, T.D., 70% efficiency of bi-state molecular machines explained by information theory, high dimensional geometry and evolutionary convergence, Nucleic Acids Research 38(18):5995–6006, 2010. Return to text.
- Ball, P., The borders of order: Do all living things exist at the edge of chaos?, New Scientist 2966:44–47, 2014. Return to text.
- Milo, R. & Phillips, R., Cell Biology: by the numbers, pp. 294–298 (June 2015 draft), Garland Science, book.bionumbers.org, accessedMay 2015. Return to text.
- Williams, A.R., Mutation: Evolution’s Engine becomes Evolution’s End, J. Creation 22(2):60–66, 2008; creation.com/evolutions-end. Return to text.
- Carter, R.W., More evidence for the reality of genetic entropy, J. Creation 28(1):16–17, 2014; creation.com/evidence-for-genetic-entropy. Return to text.
- Sanford, J.C., Genetic Entropy & the Mystery of the Genome, FMS Publications, New York, 3rd edn, 2008. Return to text.
- Marks, R.J., Behe, M.J., Dembski, W.A., Gordon, B.L. and Sanford, J.C. (Eds.), Biological Information: New Perspectives, World Scientific Publishing, Singapore, 2013. Return to text.
- Williams, A.R., Healthy genomes require recent creation, J. Creation 29(2):70–77, 2015. Return to text.
- Williams, A.R., Human genome decay and the origin of life, J. Creation 28(1):91–97, 2014; creation.com/human-genome-decay. Return to text.
- Topology/Continuity and Homeomorphisms, en.wikibooks.org, accessed August 2015. Return to text.
- Hopf fibration, wikipedia.org, accessed August 2015. Return to text.





Readers’ comments
Comments are automatically closed 14 days after publication.