

Journal of Creation 21(1):43–47, April 2007
Browse our latest digital issue Subscribe
From ape to man via genetic meltdown: a theory in crisis
A review of Genetic Entropy & The Mystery of the Genome by John C. Sanford,
Ivan Press, Lima, New York, 2005

I write this review with very mixed feelings. On the one hand, for the first time some key data are being divulged which we need to include in our models, and which honest thinkers who question evolutionist theory need to digest. But I have a problem. In the Prologue professor Sanford wrote, ‘I knew I would be at odds with the most “sacred cow” of modern academia. Among other things, it might even result in my expulsion from the academic world.’ I know John personally and treasure his intelligence and integrity. In further drawing attention to his book, I may be contributing to having his ties to academia severed, a world to which he has such strong emotional ties and to which he has made so many contributions. I know academics and journalists who have already lost their jobs for questioning Darwinian theory.
He is not exaggerating. I myself have also had my experiences in this matter.
‘I started to realize (again with trepidation), that I might be offending a lot of people’s religion,’ he confides early on. How correct he is. I recently discussed the issue of life’s origins with a dear friend I’ve worked together with for years. He brought up three arguments contra creation which I easily answered on strictly scientific terms. Suddenly he leaped to his feet. Trembling with rage he pointed a finger at me, and yelled that what I was doing was dangerous! The fundamentalists in America are dangerous! They are fighting against tolerance! They refuse to accept science! They are irrational and have no facts!
Dr Sanford is an applied geneticist semi-retired from Cornell University and now with the Institute of Creation Research. He is also the inventor of the ‘gene gun’, widely used in the genetic modification of crops. In this book the reader is confronted with compelling reasons to reject the claim that mutations plus natural selection have led to the marvels found in nature.
Many scientists do not believe man is merely the product of random mutations plus natural selection, what Sanford calls the Primary Axiom. One line of reasoning, that of irreducible complexity, has been very capably championed by professor Behe:1 molecular machines require many complex components, the absence of only one rendering that entity non-functional. Evolutionary processes cannot be expected to provide the necessary building blocks.
Others have argued that the high fidelity of DNA replication leads to very low rates of mutation. Developing humans from an ape-like forefather would just take too long. In a much cited paper, Drake has estimated2 that the rate of spontaneous mutations for humans is about 5 x10–11 nucleotides per generation. In some 6 million years from a claimed split from the chimpanzee lineage, no humans could be generated if this is true.
Sanford was a practising evolutionist and at heart a eugenicist (p. 116), who ‘gradually realized that the seemingly “great and unassailable fortress” which has been built up around the Primary Axiom is really a house of cards. … Its apparent invincibility derives largely from bluster, smoke, and mirrors’ (Prologue). But we will learn that evolutionary theory fails on grounds most people did not suspect.
Mutations are bad
Sanford forces us to recognize clearly that the relentless net effect of random mutations is degradation or complete destruction of function. After decades of research, if even one mutation out of a million really unambiguously created new information (apart from fine-tuning), we would all have heard about it by now (p. 17). This is to be distinguished from certain changes in for example bacteria (p. 19), which merely fine-tune a component of a system already in place. The changes typically involve modification of one or two nucleotides, and in huge bacterial populations these are usually already present, a solution waiting for the precise niche. In other words, ‘When we use a rheostat to dim a light, we are not creating a new circuit, nor are we in any way creating new information’ (p. 19).
Mutagens have been used for years in plant breeding, creating billions of mutation events: mostly small, sterile, sick, deformed and aberrant plants (p. 25). One improvement, low phytate corn, was caused by mutations which damaged the metabolism of phytic acid, making hungry cows happy, but hardly explaining the origin of this biochemical process (p. 25). ‘However, from all this effort, almost no meaningful crop improvement resulted. The effort was for the most part an enormous failure, and was almost entirely abandoned’ (p. 25).
Indeed, no one is suggesting replacing incubators with X-ray machines to help evolution along. On the contrary, health policies are in place aimed at reducing or minimizing mutations (p. 15).
Disastrously high mutational rates
Now Sanford provides a key fact, inimical to evolutionary theory, but fully consistent with the Second Law of Thermodynamics. The genetics community now accepts that point mutations in human reproductive cells are in the range of at least 100–300 per individual each generation (p. 34). In fact, additional kinds of mutations, such as deletions, insertions, duplications, translocations, inversions, micro-satellite mutations and all mitochondrial mutations exacerbate the situation. Mitochondrial mutations alone would add about another mutation per individual each generation within the reproductive cell line, and macro-mutations can generate more sequence divergence than all point mutations combined. The overall contributions imply more than 1,000 nucleotide changes in every person, every generation (p. 37).
Using the unrealistic lower bound of 100 mutations, and assuming 97% of the genome has no function, implies three new relevant mutations per individual each generation are generated (p. 34). Before someone attempts to shrug off these new findings, let us evaluate whether it is true that only 3% of the human genome is relevant. If the percent is twice as high, then we would double the proportion at risk through mutations.
Junk DNA or masterpiece?
Driven by an incorrect model, genomes are generally characterized as chaotic and full of meaningless evolutionary relics. The irony is that the more advanced the organism, the more so-called ‘junk DNA’ is claimed to be present (p. 37). Perhaps we should be exposing our babies to radioactivity after all?! Biochemists discover ever more complex metabolic networks, with elaborate regulatory schemes to provide feedback inhibition or acceleration. The genome is full of countless loops and branches—like a computer program using analogue and Boolean logic. It has genes that regulate genes that regulate genes, able to set in motion complex cascades of events (p. 3).
But the fact that research is steadily decreasing the proportion of supposed non-functional DNA has not been properly integrated into evolutionist thinking. ‘In just a few years, many geneticists have shifted from believing that less than 3% of the total genome is functional, to believing that more than 30% is functional—and that fraction is still growing’ (p. 21). Seriously now, when we examine organisms, such as dolphins, swallows or humans, do we get the impression of final products driven by a chaotic information processing system? In any event, in our thinking we need to start getting used to the fact that over 30 new genetically relevant, function-altering mutations occur per individual each generation.
Unity of complexity
Reductionist, materialistic thinking prevents more effective reasoning constructs from being developed. If we could understand to the finest detail the properties of all atoms in a computer we’d still fail to grasp the logic of algorithms programmed to solve a mathematical problem. We would not even suspect its existence. None of the individual components of an airplane can fly, but the integrated unity can. The purpose of a back-up in-flight computer may appear to be ‘parasitic junk’, especially if we limit our analysis to the material properties of the atoms it is constructed with. When it is to be brought into action, why and in response to what circumstances, would not be discerned by researching individual characteristics such as atomic vibrations and molecular rotations and bond strengths.
Before we assume that the information in the genome used to generate mature organisms is mostly junk, we would be wise to examine the final morphological product with more humility.
Good and bad mutations inseparable
Are mutations really causing all that much damage? Many Hollywood stars (and my wife!) sure seem awfully attractive. Since interchange of the genes provided from the father and the mother occurs, might this not provide a means of avoiding passing on defective genes? Might not ‘bad’ sperms and eggs lead to defective offspring which simply don’t survive, leaving many ‘good’ versions in the population? Well, unfortunately not. A huge number of mutations are added to the germline of every baby born, and these are spread throughout the various chromosomes. Human nucleotides exist in large linked clusters or blocks, ranging in size from 10,000 to a million, inherited in toto, and never break apart (p. 55, 81). A desirable trait will be accompanied by an undesirable trait, within the same individual (p. 79).
Therefore, within any physical linkage unit, on average, thousands of deleterious mutations would accumulate before a beneficial mutation would arise (p. 82). All of the individual 100,000–200,000 linkage blocks in genomes are deteriorating.
Furthermore, recombination appears to be primarily between genes rather than randomly between nucleotides. This means that an inferior gene is doomed to remain in that lineage, unless a back-mutation occurs, which is vanishingly unlikely. This means that the good mutations and the bad mutations cannot be separated, another example of the one-way direction of degradation known as ‘Müller’s ratchet’.
Being now clearly persuaded that the net effect of mutations will be loss of information-guided functionality, we are ready to digest another insight. Tragic as a devastating mutation may be to the affected and family, the effects of this ‘curse’ would be limited to the victim if no offspring survive. But for the population as a whole, the major damage turns out not to be the severe mutations.
Near neutrals
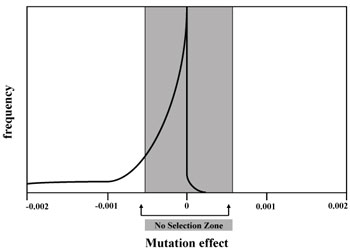
The majority of deleterious mutations have individually a negligible effect on viability of the organism. This is especially true if the ‘competitors’ are also accumulating non-deadly but nevertheless undesirable mutations. This is like the rusting of a car, one iron atom at a time (p. 72). Even one extra unnecessary nucleotide is slightly deleterious—as it slows cell replication and wastes energy (p. 21).
This issue has been mostly ignored in the literature. Mutations in the ‘near-neutral box’ (figure 1) are redefined as being completely neutral, and so dismissed. It is then claimed that more severe mutations to the left of the near-neutral box can be entirely eliminated by natural selection (p. 23). I supposed that if we are talking about a very small number of mutations this would be to a first approximation reasonable. But the accumulation of dozens or hundreds of such mutations every generation presents a totally different picture.
Incidentally, we must remember that essentially all hypothetical beneficial mutations also fall within Kimura’s ‘effectively neutral’ zone (p. 24). Therefore, positive selection would also be too weak to have an effect!
It would be desirable if natural selection could remove at least some damaging mutations. In fact, this remains our last hope to avoid a fitness meltdown. Before abandoning hope, we need to consider natural selection carefully.
Natural selection is ineffective
The same environmental factor is unable to severely penalize different deleterious mutations. It is not realistic to invoke strongly negative selection to quickly eliminate a large number of unrelated mutations. As the number of minor mutations increases, each mutation becomes noise for the others (pp. 77, 78).
Now, in a laboratory one can intelligently favour natural variability to accentuate some chosen trait (p. 98). This requires carefully crafting the external environment (nutrition, temperature, natural enemies, etc.) to minimize mutational noise. Nevertheless, no one has ever claimed to have created brand new functions not already coded for on the genome in this manner. And inevitably the organisms fine-tuned in the laboratory for a single trait are less viable long-term, living freely in nature where all natural ranges of environmental challenges occur. It is possible to optimize things such as the amount of sugar a beet produces, as long as this plant is later protected from full competition with the original stock. The changes may be in man’s interest, but at the price of the organism’s natural fitness (e.g. the large sugar production might result from a mutation damaging its control mechanism so it over-produces; in the wild, this could not compete because it is wasting valuable resources).
Outside of the laboratory the matter is much worse. There is no intelligent guidance. The judge is also nearly blind (p. 7). There is a very long chain of events separating the direct effects of a genetic change and the consequences for the whole organism level. There is a logarithmic dilution at each step, a huge loss of cause-effect resolution and correspondence. ‘It is like measuring the impact of a butterfly’s stroke—on a hurricane system which is a thousand miles away’ (p. 49). ‘It is a little like trying to select for a specific soldier, based upon the performance of his army’ (p. 49).
The literature is full of statements and abstruse computer programs claiming natural selection can perform near miracles.3–5 But after 25 years of searching, I have yet to find an analogy or computer model backing up this claim which has any biological relevance. Generally it is enough to simply ask what kind of organism would be suitable to check and perhaps calibrate the claims against, to reveal the irrelevance. Sanford offers an illustration of how natural selection really works, which reflects formally the issues involved very realistically, which I will modify to maximize correspondence to how selection really works in nature (p. 50).
Let’s imagine a new method for improving biochemistry textbooks. A few students are randomly selected who will get a biochemistry textbook each semester during the next four years, whether or not they take a biochemistry course. Each new book will have 100 random changes in the letters. Those receiving the textbook are forced to read it (whether they take the biochemistry course or not). Different teachers assign grades to all courses taken by all students across the country each semester (whether they received the biochemistry textbook or not). The correlation between true ability and each grade (math, history, Latin … ) is weak and often wrong. At the end of the semester we compare the average grades of all students nationwide and identify from among the best students those in possession of a mutated biochemistry textbook. Each of these latter textbooks are borrowed, 100 new random changes are made, and then returned to the owner. The whole cycle of reading and grading is repeated, multiple times. Will a better textbook result in this manner? No, since there is no meaningful correlation between the small differences in textbooks and the grades. Too many other factors (‘noise’), such as home life, lack of sleep, classroom setting etc. override the effect of a few misspellings.
Any trait such as intelligence, speed or strength depends on gene characteristics and environmental factors (nutrition, training, etc.) (p. 90). For example, height is about 30% (h2 = 0.3) heritable. For complex traits such as ‘fitness’ heritability values are low (i.e. 0.004). ‘This is because total fitness combines all the different types of noise from all the different aspects of the individual’ (p. 91). Low heritability means bad genotypes are very difficult to eliminate. Survival becomes primarily a matter of luck, and not better genes:
‘If Kimura’s estimate is correct, then 99.6% of phenotypic selection for fitness will be entirely wasted. This explains why simple selection for total phenotypic fitness can result in almost no genetic gain.’ (p. 93)
Natural selection is a probabilistic matter. ‘Mother Nature’ does not compute for each member of a population a ‘total fitness value’ based upon all phenotypic traits (p. 94).
Furthermore, almost all mutations are recessive, camouflaging their presence and hindering selection against them (pp. 56, 76). Another consideration, not explicitly brought out in this book, is that key environmental factors (disease, temperature, mutation, predators, etc.) affecting survival vary over time. Strong selection must be present for a huge number of generations if fixation of a (temporarily) favourable trait throughout a population is to occur. Relaxation for just a few generations could undo this process, since selection for a different trait would then be at the expense of the preceding one.
We must recognize clearly this lack of strong correlation between a mutation (whether having a positive or negative effect) and reproductive success. It is a fact of nature, yet most people attribute incorrectly near miraculous creative powers to natural selection.
But then how could natural selection supposedly develop optimized proteins, such as enzymes, one nucleotide mutation after the other, leading to almost identical versions throughout nature?6–8 Each improved nucleotide would have to be selectable in the presence of all the other noise-causing mutations within the same linkage blocks. This cannot occur by somehow selecting for superior individuals on average—which inherently involves thousands of different genes and millions of different nucleotides (p. 117).
We conclude that evolutionary theory has a major problem. If mutation/selection cannot preserve the information already within the genome, it is even more difficult to argue that billions of slight improvements were selected gradually over time (p. 106). The matter is not merely an issue of low probabilities. Theoretically a huge number of offspring could be generated, each differing by many random mutations. Might not a lot of luck bordering on the miraculous cherry-pick out the best? Not really. Sanford explains why there are physical constraints as to what natural selection could do in the real world.
The cost of selection
The number of offspring which humans can produce is rather small. For a human population to maintain its size, about three individuals per couple would be needed. This is because not all who live go on to have children, due to personal choice, accidental death, etc. Eliminating individuals carrying bad mutations would require that additional children be born, to be sacrificed to natural selection (p. 57). ‘All selection has a biological cost—meaning that we must remove (or ‘spend’) part of the breeding population’ (p. 56). In other words, deleterious mutations in man must be kept below one mutation for every three children for flawless, 100% effective selection to be able to eliminate all the mutations and still allow the population to reproduce (p. 32).
There are several kinds of costs, all additive, which must be paid for before ‘real’ selection can be covered (p. 59).9 As mentioned above, fitness has low heritability, meaning environmental factors are much more important than genetic factors in determining who survives. This means that a very large number of additional offspring is needed, which must die due to natural selection independent of genetic causes, simply to remove non-heritable variations (p. 59). In these circumstances, having to additionally select the worse culprits which carry 100 or more mutations, every generation, is not physically possible (p. 62).
Haldane’s Dilemma

Having demonstrated conclusively that the degradation of the human genome (in the presence of such high mutations rates, preponderance of deleterious mutations and lack of huge expendable proportions of offspring) cannot be avoided, we return to what evolutionary theory claims happened. Ever more complex and sophisticated genomes are supposed to have arisen, step by step, over eons.
In the 1950s, one of the most famous population geneticists, John Burdon Sanderson Haldane, presented an observation known as ‘Haldane’s dilemma’ (p. 128): it would take (on average) 300 generations to select a single new mutation to fixation. However, his calculations were only for independent, unlinked mutations. He assumed constant and very strong selection for a single trait, which is not realistic. The interference by hundreds of random mutations was not taken into account. Even so, selection for only 1,000 specific and adjacent mutations could not happen in all putative evolutionary time. There is no way an ape-like creature could have been transformed into a human (p. 129). Man and chimp differ at roughly 150 million nucleotide positions (p. 130) and humans show remarkably little variation worldwide.
Think for yourself
Advanced education is dominated by evolutionary theory taught as established fact. But ‘are you really just a meaningless bag of molecules—the product of nothing more than random molecular mutations and reproductive filtering?’ (Prologue). This doctrine is presented as unquestioned truth, an axiom accepted by faith because many scientists present it as obviously true (p. 5). But if you come to the point where you feel that the Primary Axiom is no longer obviously true to all reasonable parties, then you must not accept it on blind faith (p. 10). At best the materialist model could be basically right, but it is absurd to continue believing that it is self-evident. At the very least, critical thought and fair discussion is required, something scorned and denigrated by the current high priests of biology.
Historically, the entire field of population genetics was developed by a small, tightly knit group of people radically committed to the Primary Axiom. They were free to explore many scenarios and adjust multiple parameters unconstrained by objective calibrations, and to optimize frameworks to appear internally consistent. Their mathematical approach was to define the unit of selection as discrete genetic units, such a gene or nucleotide, instead of whole organisms with all the contradictory influencing factors (p. 52).
‘For the most part, other biologists do not even understand their work—but accept their conclusions “by faith”’ (p. 46). The theorists’ models can be shown to never have matched biological reality to the minimal degree expected of useful models, but these men were undeniably intelligent and had an incredible aura of intellectual authority (p. 53). In many ways they deserve our admiration, since transforming any scenario, correct or not, into an appropriate mathematical formulation requires a great deal of skill. One can also admire honestly the brilliant lawyer who argues ever so cleverly against the truth in his client’s interest. How we wish they would contribute their gifts within a correct paradigm!
There is hope
Finally, professor Sanford makes it clear that no amount of human intervention can salvage the relentless degradation of our genomes. We will experience much and increasing suffering on the part of our children and grandchildren. But our Creator made the genome in the first place.
‘ … Jesus is our hope … He gave us life in the first place—so He can give us new life today. He made heaven and earth in the first place—so He can make a new heaven and earth in the future’ (p. 155).
Read this book twice. Then read it again with a highlighter. Technical aspects are easy to follow, and the specialist will benefit very much for the highly relevant references offered.
References
- Behe, M., Darwin’s Black Box: The Biochemical Challenge to Evolution, The Free Press, New York, NY, 1996. Return to text.
- Drake, J.W., Charlesworth, B., Charlesworth, D. and Crow, J.F., Rates of spontaneous mutation, Genetics 148:1,667–1,686, 1998. Return to text.
- Dawkins, R., The Blind Watchmaker, Penguin Books, London, 1986. Return to text.
- Lenski, R.E., Ofria, C., Pennock, R.T. and Adami, C, The evolutionary origin of complex features, Nature 423(6936):139–144, 2003. Return to text.
- Schneider, T.D., Evolution of biological information, Nucleic Acids Res. 28:2794–2799, 2000. Return to text.
- Truman, R. and Heisig, M., Protein families: chance or design? Journal of Creation 15(3):115–127, 2001; creation.com/protein. Return to text.
- Truman, R., The ubiquitin protein: chance or design? Journal of Creation 19(3):116–127, 2005; creation.com/ubiquitin. Return to text.
- Truman, R., Searching for needles in a haystack, Journal of Creation 20(2):90–99, 2006. Return to text.
- ReMine, W.J, Cost theory and the cost of substitution—a clarification, Journal of Creation 19(1):113–125, 2005; creation.com/cost. Return to text.



Readers’ comments
Comments are automatically closed 14 days after publication.