
Apoptosis: cell ‘death’ reveals Creation
Apoptosis* (‘programmed cell death’) is a biologically ubiquitous phenomenon that deserves to be much more widely known among non-biologists and laypeople. Put quite simply, without apoptosis, all multicellular life would be impossible. From a creationist perspective, the vast literature that exists on this subject is seen to be entirely compatible with its origin as a created process. It is genetically programmed, exquisitely regulated and exhibits a mind-blowing complexity that speaks eloquently of its design by our Creator God. This is demonstrated first by looking at the many and varied roles it serves in healthy organisms (particularly human beings). Secondly, malfunctioning apoptotic mechanisms are shown to cause many of the debilitating diseases that we are familiar with in this sin-cursed world. Not surprisingly, this is a hot research area, the 1990s seeing a literally exponential rise in new publications about apoptosis. This article reviews the main findings of this burgeoning literature and offers perspectives on the relevance of apoptosis for the creation/evolution debate.
* Terms marked with an asterisk are defined in the Glossary at the end of this article.
Click here for larger view.
Very few of the general public will have heard of apoptosis (programmed cell death; PCD). In fact, the majority of professional scientists are only vaguely aware of what it is. This is partly due to its total absence from secondary school science curricula and partly because it has remained in the domain of research biologists. Apoptosis has actually been known for some time—first described around 30 years ago. During the 1990s, the research literature saw an explosion of interest in this area, even spawning two new scientific journals,1,2 devoted solely to this topic. This was due, in no small part, to the growing realisation that, here was a phenomenon that had far-reaching implications for our understanding of human diseases such as cancer. Far from being limited to a few obscure organisms, apoptosis has turned out to be a ubiquitous biological phenomenon. It is known to occur in all multicellular organisms, from the humble nematode, Caenorhabditis elegans (where it has been much studied), to man. It now constitutes an essential part of our understanding in many diverse biological and medical disciplines. Without the cells’ ability to undergo apoptosis there would be severe dysregulation* of the maintenance and development of multicellular animals: embryogenesis*, tissue differentiation* and morphological development would not be possible; clonal selection* in the immune system could not occur; haemopoiesis (the production of blood cells) would not be properly controlled, if at all; processes such as metamorphosis, neural development and epithelial cell turnover would be disrupted. Conversely, in human beings, aberrant apoptotic mechanisms are now known to be involved in such illnesses as cancer, leukaemia, lymphoma, stroke, heart disease, rheumatoid arthritis and AIDS.
What is apoptosis?
In spite of its discovery by Carl Vogt3 as far back as 1842, research lay dormant until recent decades. An online search of the U.S. National Library of Medicine’s PubMed (which provides access to over 11 million medical citations back to the mid-1960s and additional life science journals) produced just six papers on PCD prior to 1972. The term apoptosis was coined in 1972, from the Greek apo (=off) + ptosis (= a falling or dropping off), and defined as ‘a basic biological phenomenon with wide-ranging implications in tissue kinetics’.4 There is still no consensus among scientists about how to pronounce apoptosis and both the following are equally popular: ã-pop-to’sis and ã-po-to’sis. Until quite recently, the word was not listed in major dictionaries. Stedman’s Medical Dictionary (1995) defines it as ‘a single deletion of scattered cells by fragmentation into membrane-bound particles which are phagocytosed by other cells; believed to be due to programmed cell death’. Another synonym that is sometimes used is ‘cell suicide’. For a decade, the average number of research papers about apoptosis published per year barely reached double figures. That would soon change.
An intrinsic cellular program
All cells eventually die, sooner or later. This longevity of cells varies from approximately 120 days (for red blood cells) to 120 years (pluripotent stem cells*), the current, approximate upper limit for human life span. These figures, however, depend on whether the cell is aberrant or normal (e.g. sickle-celled versus normal erythrocytes*). Even without injury or adverse conditions, cells may ‘die’. That is, an in-built cellular programme is enacted which terminates the existence of the cell after a predetermined period. This is an entirely natural, physiological process; indeed it is essential to the health of an organ or the organism as a whole (see later). The process itself is under genetic control, is very finely tuned, involves macromolecular synthesis and incorporates multiple signalling pathways. Furthermore, numerous components interact with one another, allowing many ramification points, so that the same trigger can induce different intracellular effects.5
An apoptotic pathway, once triggered, involves an energy-dependent cascade of biochemical and molecular changes in the cell. In this respect, it has similarities to the blood-clotting process6 and the complement pathways (part of the innate immune system);7 i.e. all components have to be present for functionality, an insurmountable barrier to gradualist explanations of its genesis.
Avenues of early apoptosis research
In the mid 1980s, work on the 1 mm long nematode, Caenorhabditis elegans, revealed that apoptosis was vital to its normal development. The sequential development of the 1,090 cells leading to the adult worm was well known. Researchers were able to knock out specific cells (using a laser) and examine the effects of this on the subsequent growth; conveniently, C. elegans takes just three days to mature. They were then able to elucidate the precise location and timing of 131 cells that underwent apoptosis. The activation of two genes was found to be necessary for apoptosis to occur: ced-3 (cell death gene 3) and ced-4.8 In 1992, it was reported that apoptosis in C. elegans could also be inhibited. A new gene, ced-9, was found to control the activation of ced-3 and ced-4. Only when ced-9 was inactivated was the block on apoptosis removed.9 Just months later, Bcl-2, a well-known gene involved in human B-cell lymphoma, was also shown to be able to protect against apoptosis, when transfected into C. elegans. Such results led to the belief that apoptosis was as universal a cellular phenomenon as proliferation.10
A rapidly expanding field
Since 1993, the number of articles on this subject published per year has increased by an order of magnitude. Today, scientific knowledge of the apoptotic regulatory machinery has increased enormously, with numerous genes and proteins now known to be involved. In a 1997 analysis of the impact of this research phenomenon, the authors stated ‘PCD is now one of the hottest areas in science. In fact, among the 10 most-cited scientific papers published in 1995 and cited in 1995, four concern PCD.’11 In the year 2000, more than 700 papers (on some aspect of apoptosis-regulation) were published in scientific and medical journals each month! This contrasts with approximately 65 new papers related to mitosis (cell division) each month in the same year.12
Mechanisms and their regulation
The first line proteins are those containing a so-called ‘death domain’ (DD). Some of these are trans-membrane proteins with a cell-surface receptor portion: the much-studied Fas(APO-1/CD95), the subject of well over 3,000 research articles to date, including one by this author;13 TNFR1; DR3(TRAMP/wsl-1/APO-3/LARD/AIR); and DR4(TRAIL-R1/APO-2). The many alternative names arose from the fact that previous reports of different apoptosis-related proteins turned out, with subsequent research, to be describing the same protein. In 1995, two competing groups simultaneously reported that trimerisation* of the Fas and TNFR1 ‘death receptors’ by ligand (Fas-L and TNF-α respectively) induced apoptosis.14,15
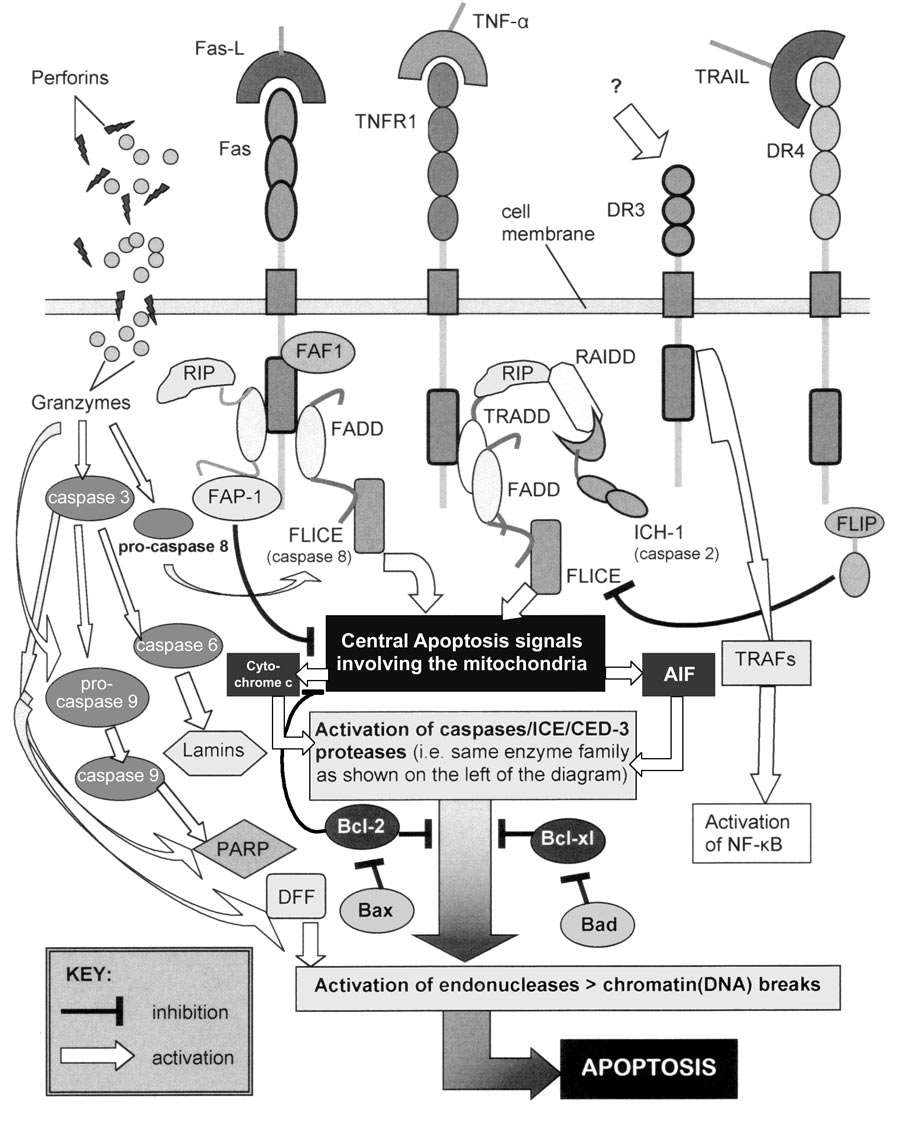
Intracellular ‘adaptor’ proteins (such as TRADD, FADD/MORT1, RIP, RAIDD/CRADD and Reaper) have death domain homologous regions* that bind to the DDs of Fas and TNFR1,16-19 thereby relaying signals to a further series of proteins, variously known as the ICE/CED-3/caspase family (of which numerous types have been described20 ). These caspases* are basically protease enzymes and have been dubbed the apoptosis ‘executioners’21 because their activation seems to be a universal occurrence in apoptosis, although alternate/redundant apoptotic pathways may exist. The mitochondria* play a central role in this caspase activation by releasing molecules into the cytoplasm that help execute apoptosis, chiefly cytochrome c,22,23 but also AIF (apoptosis-inducing factor).24
The picture of apoptosis activation/regulation is further complicated by yet other functional protein families, the members of which variously act as promoters or inhibitors of the process. For example Bcl-2, mentioned earlier, gives its name to a family of proteins that regulate the caspases.25,26 Bcl-2, Bcl-xL, A1, Mcl-1, Ced-9 and BHRF1 (Epstein-Barr virus) act as inhibitors, whereas Bax, Bcl-xS, Bad, Bak and Bik/Nbk, act as promoters of apoptosis. If the sheer numbers of different components involved in apoptosis and its regulation were not startling enough—and there are many more than mentioned above (such as p53,27 Rb,28 perforins,29 granzymes30 and ceramide31 )—their multifarious interactions (additive, synergistic, antagonistic, promotor, inhibitive, etc.) result in a level of complexity that is truly awesome (Figure 1). This, of course, is powerful testimony to our omniscient Creator God, who designed these cellular systems so perfectly in the beginning.
What apoptosis is not
It is crucial, at this juncture, to state that apoptosis was not an unwelcome intrusion into the perfect world that God created (Genesis 1:31). Superficially, this assertion may seem incongruous with the descriptions ‘programmed cell death’ and ‘cell suicide’ but these terms are simply anthropomorphisms—human attempts to give a name to a process, whereby cells are removed/deleted from the body.
The Bible insists that death, the ‘last enemy’ (1 Corinthians 15:26), was a consequence of Adam’s sin (Genesis 2:17; 3:17–19; Romans 5:12, 14, 17, 21; 1 Corinthians 15:21–22). Death, disease and suffering are familiar symptoms of our fallen world. As has been well documented by creationists, the physical death that was inaugurated by the Fall referred to ‘nephesh life’.32–35 In fact, the presence of apoptosis throughout the living world is, as we shall see, a powerful testimony to a Super-Intelligent Designer, our Creator God Himself. It is universally true that certain diseases (with the associated suffering and death) occur precisely because of errors in apoptotic mechanisms, as we shall see later. Disease does not result from the occurrence of apoptosis per se.
It is because I want to avoid the negative connotations of death, that I choose to use the term ‘apoptosis’ whenever possible. It is more appropriate to view apoptosis as a cellular response to stimuli, as this implies the existence of target cells and the optimum levels of specific stimuli.
Apoptosis versus necrosis
** An injurious agent may activate at least four major ‘pathways’ leading to a loss of membrane integrity (of particular relevance to necrotic death): 1) Membrane phospholipid degradation; 2) Production of amphipathic lipids; 3) Damage to cytoskeleton; 4) Generation of toxic oxygen species and free radicals.
***The contrasting changes in cell volume for apoptosis and necrosis are both osmotic effects. For instance, in the case of necrosis, the swelling of the cytoplasm is probably due to an increased ‘osmotic load’, resulting from the accumulation of waste products of metabolised xenotoxin or of the toxin itself.
Click here for larger view.
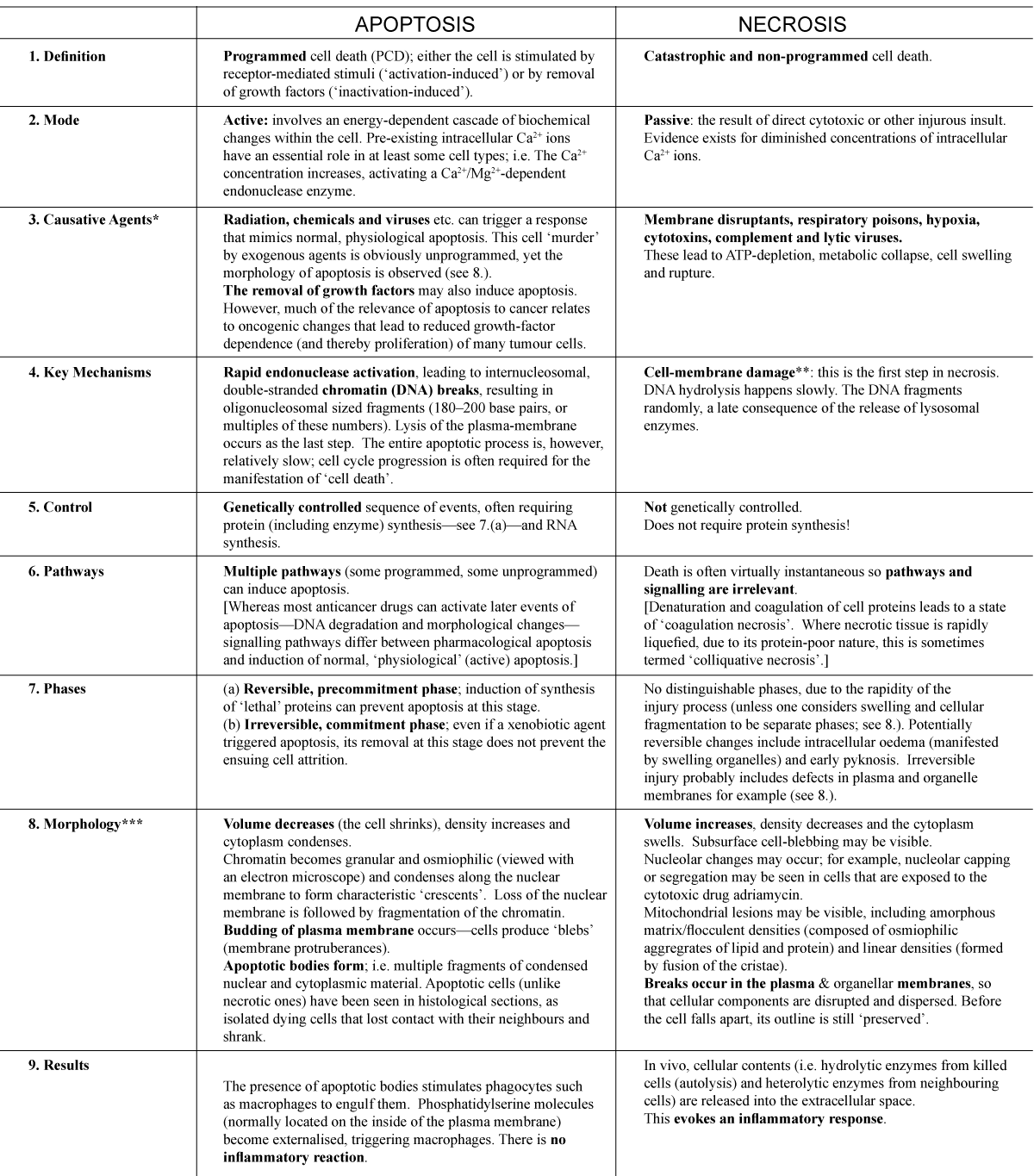
Necrosis* can be thought of as the second fundamental ‘mode’ of cellular death and occurs when cells are injured in such a way that the normal apoptotic mechanisms are circumvented. Unlike necrosis (see below), apoptosis is accompanied by characteristic morphological changes that invariably include a loss of the normal membrane asymmetry*, reduced cell volume, chromatin* condensation and fragmentation of the chromosomal DNA (Figure 2). If/when a cell is deleted (‘dies’) by apoptosis, all traces of it are efficiently removed by other ‘dedicated’ cells, with a total absence of inflammation*. This culminating phagocytosis* of the residual nuclear and cytoplasmic components (so-called ‘apoptotic bodies’) by macrophages* is part of the natural cellular economy; indeed, it facilitates the reprocessing and re-utilisation of materials. As we saw earlier, apoptosis actually requires genetic programming and therefore information input36 and often involves protein and RNA* synthesis. In stark contrast, necrosis is chaotic and catastrophic; i.e. a cellular example of increased disorder, in perfect accord with the Second Law of Thermodynamics. Whereas apoptosis is absolutely essential to increases in order and complexity of multicellular organisms, necrosis results in the accumulating detritus of large numbers of dead/dying cells, which cause problems for the organism unless it is swiftly dealt with. Necrotic cells are therefore indicative of a world where decay has set in, a consequence of Adam’s sin. Those readers who are interested in a more detailed comparison of apoptosis and necrosis are referred to Table1. While the precise details of apoptotic morphology and molecular biology may vary for different cell types, the same goal of physiological cell deletion is accomplished in each case.
Apoptosis versus mitosis
It is now realised that the processes of apoptosis and mitosis are really ‘two sides of the same coin’. Mitosis, as all biology students are taught, is cell multiplication, leading to tissue growth. Apoptosis is cell deletion, leading to tissue shrinkage locally. The intentional rhyming of apoptosis with mitosis shows that there is normally a balance between cell ‘death’ and cell birth respectively. If apoptosis is a universal phenomenon among eukaryotic*, multicellular organisms, why did it escape the attention of biologists for so long and why is it still absent from most dictionaries and school science curricula? Lockshin offers the following reflections:
‘Mitosis is a relatively short and evanescent event, but once it has occurred there are traces of its passing: a single-celled egg becomes two, four, and eight … . Apoptosis is far more furtive. Cells simply vanish, usually in less than one hour.’50
It is this transient nature of the apoptotic signature that seems to have kept its true relevance hidden from the scientific community until relatively recently. As we shall now consider, apoptosis is literally vital to multicellular life.
Domains for the occurrence of apoptosis
It is beyond the scope of this paper to elaborate on the various roles of apoptosis in detail, but the following examples (in healthy animals and humans) illustrate how ubiquitous apoptosis is in biology:
- Apoptosis is essential during vertebrate embryological development in order to correctly sculpt the multifarious tissues and organs that are forming. The classical example is that of the developing limbs. These initially form as ‘buds’, the digits (e.g. fingers) forming later on, by virtue of apoptosis of the cells in the interdigital areas.51,52
- Apoptosis plays a pivotal role during both T-and B-lymphocyte maturation.53 For instance, the elimination of mature T-cell clones in the peripheral blood seems to be a mechanism for establishing tolerance to self antigens.54 Also, apoptosis may be activated during normal B-cell ontogeny*.55,56
- The human eye is a classic example of so-called ‘immunologic privilege’ by virtue of the fact that the exposed eye surfaces express the Fas-L(CD95L) molecule. If the eye is attacked by virus, a massive inflammatory response ensues, but no long-term damage to the eye occurs because the Fas-L triggers apoptosis in the infiltrating cells (neutrophils* and T cells), which are all removed in a few hours.57 This is obviously essential to healthy vision!
- Apoptosis provides a safe, disposal mechanism for neutrophil granulocytes at inflamed sites. Human neutrophils ingest foreign microbes at sites of injury, but their prolonged survival (once the job is done) would cause chronic tissue damage. It is now thought that they actively generate ROIs (reactive oxygen intermediates) in order to mediate their own speedy demise.58
- Apoptosis serves a vital role in controlling the number of germ cells in the testis59 and selectively eliminates cells with high proliferative activity, that acquire irreparable genetic abnormalities; e.g. testicular temperature rise causes more mutations in prospective gametes but heating to higher temperatures results in aspermia*.60 That is, apoptosis is very important in limiting teratogenesis (the production of neonatal abnormalities).
- In the ovary, a vast amount of apoptosis occurs in the germ-line, throughout the later stages of pre-natal development and on into post-natal life. One example is that it sets the absolute number of oocytes (eggs) that are available for development and ovulation in adult life.61

Figure 2. Schematic diagram of the morphological features of apoptosis and necrosis (adapted and redrawn from103).
Click here for larger view. - In non-pregnant women, the cyclic fluctuations of the menstrual cycle hormones determine the cellular fate (proliferation, differentiation, or apoptosis) of both mammary gland epithelium*62 and the uterine endometrium*.63 Menstruation itself is brought about by apoptosis of a specific endometrial cell population. In post-pregnancy women and mammals generally, regular suckling ensures that lactation continues. Conversely, at weaning, the gradual or abrupt cessation of milking of the mammary gland results in its gradual or rapid involution respectively. This involution (leading to milk stasis) is due to a net loss of glandular mammary tissue by apoptosis.64
- Human breast milk contains the common protein alpha-lactalbumin which, in its multimeric* form, has been reported to have potent apoptosis-inducing effects in tumour cell lines, but much less effect on a variety of normal cell types.65 I have personally confirmed these results in a series of experiments on fresh lymphocytes from healthy volunteers and from patients with chronic lymphocytic leukaemia.66 The original discovery was serendipitous67 and the precise function of this capability of breast milk is not fully understood. Perhaps the induction of apoptosis in transformed, but not mature, epithelial cells helps direct the normal growth of neonatal mucosal* epithelium and helps prevent neoplasia*.65
- In the nematode Caenorhabditis elegans, the morphological development is very precisely governed by which cells divide and which cells undergo apoptosis at specific stages.8,9
- Metamorphosis brings about pronounced morphogenetic changes in a short time as organisms change from the larval to adult stages. Apoptosis plays a central role in this process. Metamorphosis is well-known in numerous insect species, as well as certain vertebrate organisms. For instance, metamorphosis involves the complete remodelling of virtually every tissue/organ as tadpoles transform into frogs. Most organs, if they are not completely reabsorbed, at least undergo apoptotic deletion of certain cells.68

It is not really possible to do justice to the role of apoptosis in each of the above but the interested reader is referred to the references. A recommended recent book that covers some of this material in more detail is When Cells Die. In the preface, the editors make the following, interesting comment:
‘Throughout the book, the clearestconsensus is thatan organism uses cell death in a very positive way—to sculpt its development, to arrange for rapid expansion and subsequent contraction of a cell population in the immune and reproductive systems, and to defend itself by destroying cells that have been infected or attacked’ [emphasis added].69
A biblical perspective on the purpose of apoptosis
It is clear from the foregoing, that apoptosis is an essential physiological mechanism and as such, would have been present in the pre-Fall world. However, as some of the above examples show, some apoptotic pathways are also triggered as a toxicological reaction. Thus, they are vital to an organism’s health (even survival) during exposure to pathogenic* microorganisms or environmental stress. Apoptosis can be viewed as the ultimate mechanism for maintaining phenotypic fidelity* in multicellular organisms.70
From a biblical perspective, the capacity for deployment of these abortive cell-processes (as a ‘stress-response’) was built into the genetic potential of all organisms by our Omniscient Creator. After the Fall (and the resulting curse on all Creation), the role for apoptosis must have diversified substantially. Conditions in the radically altered, post-Flood world would, no doubt, have added to the ‘work-load’ of the originally created apoptotic mechanisms. Obviously, with the passage of time, entropy was increasingly manifest by these originally perfect systems and we see the results today, when defective apoptotic responses sometimes lead to disease (see below).
It is of interest to creationists that failure of normal apoptosis has been proposed as a possible means to facilitate the aging process.70 Various experimental studies of the longevity of human lymphocytes have correlated apoptosis with aging.71 Perhaps degeneration of apoptotic control increases with time so that it has a rate-limiting effect on the aging process. This has obvious implications for our thinking about the antediluvian long lifespans. Bergman has recently discussed this, with respect to the telomeres that protect the ends of normal cellular chromosomes.72 These telomeres shorten with each cell division, eventually triggering senescence* and apoptosis. Bergman speculates that:
‘At some point in history human longevity could have changed as a result of some alteration of the telomerase system.’73
When apoptosis goes awry
The perfect functioning of an apoptotic mechanism depends on the genetic information being uncorrupted, as it was at Creation. Therefore, any loss of information, such as occurs when a gene that codes for a protein in an apoptotic cascade mutates, will almost certainly have drastic consequences. Indeed, the sensitivity of these mechanisms to the slightest change in the configuration of a single protein component, is a powerful argument against neo-Darwinism. It is truly inconceivable that random changes to gene-encoded information for these pathways could ever produce an improvement. Rather, apoptotic pathways have all the hallmarks of irreducible complexity74 and attempting to construct them in a step-wise fashion, whilst maintaining functionality at each step, would be futile in the extreme.
Dysregulation (too much or too little) of apoptosis can cause a wide spectrum of defects:
- Cancer. Defects of normal apoptotic processes have been discovered in many forms of cancer.75 Normal cells maintain a balance between the rates of mitosis and apoptosis. However, apoptotic failure of a cell that has sustained one or more somatic mutations results in an immortalised ‘cellular anarchist’ (neoplastic* cell). Solid tumours (cancers) or an uncontrolled proliferation of haematopoietic cells (leukaemia and lymphoma) are the inevitable consequence. Since many genes regulate apoptosis, it is no surprise that defective genes have been noted in many cancers (so-called oncogenes), such as p5376 and bcl-2.77,78 Over-expression of Bcl-2 protein, as a result of bcl-2 oncogenesis, confers resistance to the neoplastic cells so that they are much less susceptible to chemotherapeutic drugs and radiotherapy.79
- AIDS. Apoptosis research has thrown light onto the causes of this immune deficiency syndrome. Among other things, it seems that expression of Fas(APO1/CD95), a ‘cell death’ receptor, is enhanced in individuals infected with HIV (human immunodeficiency virus), contributing to/causing an increase in the apoptotic rate of CD4+ T-lymphocytes.80 This is however, somewhat controversial among AIDS researchers.81,82
- Alzheimer’s disease (AD). Individuals with AD suffer premature or excessive neuronal cell loss in the brain during the aging process, together with other pathological effects (the formation of plaques, gliosis* and neurofibrillary tangles*). Compromised mitochondria may release a significant amount of calcium ions into the cytoplasm, so stimulating the caspases and DNases involved in apoptosis.83 However, there is some ambiguity regarding the role of caspase-dependent neuronal apoptosis in AD due to contradictory experimental observations and a lack of convincingly apoptotic neurons in AD brains.84 Other AD research indicates that the apoptotic signal may not reach the terminal caspases.85 Thus, while apoptosis certainly seems to be implicated in AD, a causative role is questionable.
- Rheumatoid arthritis (RA). RA is an autoimmune disorder in which the body attacks its own cartilage in the synovial joint linings, leading to inflammation, painful swelling and eventually loss of joint function. It has been shown that chondrocytes (cartilage cells) of articulating bone surfaces are more prone to apoptosis in RA patients.86 This was recently found to be strongly associated with expression of pro-apoptotic proteins such as Fas(APO1/CD95) Fas-L and p53.87 The cause of RA is essentially due to multi-level aberrations that lead to defective apoptosis or hyperapoptosis.88
- Embryonic lethality. Mice are widely used as disease models in medical research. There are many reports in the scientific literature of murine embryos dying due to excessive apoptosis, either because a particular protein is overexpressed, or because it is deficient. For example, embryos die in utero around mid-gestation if they are deficient in cytochrome c89 or the nuclear factor kappaB.90 In both of the latter cases, the authors demonstrate that the protein deficiency renders the mice susceptible to apoptotic signals, mediated by TNF-α/TNFR1 signalling.
- Eye problems. Apoptosis has critical (and contrasting) roles in the various ocular tissues (cornea, lens and retina) and extraocular tissues (e.g. optic nerve) that contribute to vision. We saw earlier that it contributes to immunologic privilege in the healthy eye. It may also initiate healing of eye wounds but malfunctioning apoptotic pathways are also associated with opthalmological disease.91
In every case, small perturbations of apoptotic mechanisms (often resulting from a single somatic mutation) have debilitating, even lethal effects, belying ideas of genetic gradualism and corroborating Behe’s thesis of ‘irreducible complexity’.74 It is clear that impairment of apoptosis compromises the body’s ability to effectively eliminate damaged or mutated cells, which would affect the organism’s survival if they lived on.
The therapeutic potential of manipulating apoptosis
Leading researchers in this field recently stated that:
‘Programmed cell death and apoptosis are very important aspects of a healthy life, and our access to manipulation of it will have vast consequences in many fields of medicine and agriculture.’92

Click here for larger view.
Restoration of the apoptotic response would be beneficial in many cases. This is a particularly fruitful avenue of research with respect to treating many cancers. Clearly, the very existence of cancer is testimony to the ongoing genetic degeneration that started with the Edenic curse.93 In cancer and haematological* neoplasms, restoration of the apoptotic response to therapeutics would help to solve the longstanding problem of multi-drug resistance of malignant cells.94 For example, in healthy people, Fas(APO-1/CD95)-mediated apoptosis is thought to be responsible for the removal of anergic*, autoreactive B-lymphocytes from the peripheral blood circulation; i.e. part of a normally functioning immune system. Once activated, mature B-cells acquire surface Fas-expression and concomitant sensitivity to apoptosis, induced by Fas-ligand on Th1 (T-helper) cells. However, malignant cells from patients with B-cell chronic lymphocytic leukaemia or mantle cell lymphoma (see Figure 3) usually lack surface Fas expression, rendering them resistant to Fas-mediated apoptosis. I recently co-authored the report of a study, in which we attempted to re-sensitize such neoplastic lymphocytes to apoptosis using the cytokine, interleukin‑2.97
Inhibition of inappropriate apoptosis could be beneficial in other cases. This is particularly the case, where cellular degeneration results from a disease process that causes too much apoptosis. Approaches aimed at reducing apoptosis are an active research area for scientists studying viral pathogenesis (e.g. HIV infection),80 neuronal degeneration (e.g. Alzheimer’s disease)83 and rheumatoid arthritis,87,88 for example.
Conclusions: apoptosis from a creation/evolution perspective
Faced with the growing realisation that apoptosis is one of the fundamentally important biological processes, some evolutionists are beginning to grapple with such crucial questions as how it originated and how it was selected for, according to Darwinian, selectionist dogma. Comments such as the following are predictable:
‘Many of the genes that control apoptosis are conserved throughout evolution from mammals to nematodes, flies, and viruses.’98
Similarly,
‘The preservation of a process throughout evolution indicates that the process is fundamental and too important to be modified.’99
Clearly however, this type of remark merely begs the question of how apoptosis is supposed to have evolved in the first place.100 Another paper by this author reviews the few evolutionists’ attempts to explain apoptotic origins and argues that the very existence of apoptosis actually falsifies evolution.101 Contrary to the impression given, the great diversity exhibited by apoptotic pathways, notwithstanding fundamental similarities, is a hallmark of Creative Design.102 Apoptosis is widely distributed among disparate species and tissues. Also, whereas a single mechanism of action would be predicted by reductionists, we observe multiple effectors.
So, just how far back did apoptosis originate in an evolutionary world? According to the evolutionary paradigm, injurious agents (e.g. irradiation, cosmic radiation, free radicals, hypoxia/hyperoxia*, environmental chemicals) and hyperthermic conditions have characterised the early Earth’s history at one time or another. Thus, a sophisticated apoptotic response would have been especially necessary at the very stage in Earth’s history when multicellular, eukaryotic cells are supposed to have evolved (during the late Precambrian)! Even today, apoptosis serves an essential role in terms of ‘cellular altruism’. It helps to ensure that an organism’s genetic integrity is not compromised, by removing some somatic cells that have sustained irreparable, genetic mutations. Crucially, apoptosis also helps to maintain a species’ genetic integrity, by eliminating aberrant germ cells that would otherwise carry intact but faulty genes into the next generation. Thus, on the hypothetical early Earth of the evolutionist, fully-functional apoptosis would have been much more important than it is today!
In summary, a creationist perspective is cognizant that apoptosis:
- is as vital and ubiquitous to life as mitosis;
- is programmed and tightly regulated—speaking of a Programmer who put the information into cells originally;
- is irreducibly complex—it only takes one part of the mechanism to malfunction and the whole process is badly affected, especially illustrated by cancer;
- shouts Design (Romans 1:20);
- is a Created mechanism—part of the ‘very good’ declaration of God (Genesis 1:31).
Acknowledgements
I am sincerely grateful to the Lord for planting the idea for this article in the first place, to Carl Wieland for his encouragement, and to the two reviewers, whose comments have helped me to enhance the paper.
Glossary |
|
| anergic | in a state of being unable to react to antigenic (immunogenic, allergenic) substances. |
| apoptosis | an active process (requiring energy) involving the programmed deletion of scattered cells by fragmentation into membrane-bound bodies which are phagocytosed by other cells. No inflammatory response occurs. |
| aspermia | a total absence of spermatozoa (sperm cells) in an ejaculate. |
| caspases | a group of enzymes which are particularly involved in the transduction of signals for apoptosis. |
| chromatin | the genetic material of the nucleus, made up of DNA and protein. During mitosis, the chromatin condenses into chromosomes. |
| clonalselection | the specific recognition of foreign protein (antigen) by a small proportion of the body’s circulating lymphocytes, followed by the rapid expansion of this lymphocyte clone for specific antibody production and immunologic memory. |
| differentiation (cells/tissues) | the development of morphology and/or functions that were not part of the original cells/tissues; occurs to bring about greater specialisation. |
| dysregulation | dys is a prefix meaning mis- or un-, so this refers to abnormal regulation. |
| embryogenesis | the formation of the characteristic configuration of an embryo’s body. |
| endometrium | the mucous membrane that forms the uterus lining, consisting of columnar epithelium and glands; its structure and thickness vary in accordance with the menstrual cycle. |
| epithelium | the cellular layer that covers all free body surfaces, e.g. cutaneous (skin) and mucous layers. |
| erythrocyte | a red blood cell; lacking a nucleus and specialised for oxygen transport. |
| eukaryotic/eukaryote | literally means those organisms with ‘good nuclei’ in their cells (i.e. all animals, plants and fungi); the nucleus is membrane-bound and its DNA is associated with proteins to form chromosomes. The cytoplasm also contains many organelles (e.g. mitochondria) that are absent from prokaryotes (e.g. bacteria and ‘blue-green algae’). |
| gliosis | an overgrowth of the astrocytes (star-shaped neural cells) in an area of brain or spinal cord damage. |
| haematological | pertaining to the blood and blood-forming tissues. |
| homologous regions | similar/equivalent amino-acid sequence; an evolutionist would interpret this as evidence of their common molecular origin in a putative ancestor. |
| hypoxia/hyperoxia | respectively, too little oxygen or too much oxygen; i.e. oxygen tension (a key physical property for living organisms) changes with pressures that differ from 1 atmosphere. |
| inflammation | essentially, a pathological process that results from some sort of injury, usually involving redness, heat, swelling and pain. |
| macrophages | large, long-lived, phagocytic cells that are a major part of the body’s immune defences; their morphology varies a great deal, but all are derived from monocytic stem cells of the bone marrow. |
| membrane asymmetry | the phospholipid (‘fatty’) membranes of living cells have many associated protein molecules, some spanning the entire membrane, some located towards the outside of the cell and some inside; basically a non-symmetrical arrangement. |
| mitochondria | subcellular organelles; the power houses found in all eukaryotic cells that provide the energy (in the form of ATP) to drive all cellular reactions. |
| mucosal | pertaining to a mucous membrane (tissue layer); various types of mucosa line the body’s tubular cavities, such as the stomach, intestines, trachea, uterus etc. They are composed of epithelium and mucous glands. |
| multimeric | a grouped arrangement of several identical molecules, e.g. a trimer consists of three molecules. |
| necrosis | cell death that results from injury or a pathological condition; it is a passive, chaotic process. |
| neoplasia/neoplastic | a pathological process that results in neoplastic tissue; i.e. abnormal cells that proliferate uncontrollably and generally more rapidly. |
| neurofibrillarytangles | the accumulations of disorganized filamentous tissue between nerve cells; patients with Alzheimer’s disease exhibit these in the hippocampus and cerebral cortex (brain regions). |
| neutrophils | a class of granulocytes, mature white blood cells with a lobed nucleus and a granular cytoplasm; so-called because their nuclear material does not show a particular affinity for either the acidic or basic stains that are commonly used in haematological laboratories. |
| ontogeny | the history of an organism’s development. |
| pathogenic | having the capacity to cause disease. |
| phagocytosis | literally means ‘cell-eating’; phagocytes are cells which ingest (then digest) foreign particles, bacteria, necrotic tissue, etc. |
| phenotypic fidelity | describes how faithfully the observable characteristics (manifestation of the genetic makeup) of an organism are conserved. |
| pluripotentstem cells | non-specialised, primordial cells that can differentiate into a wide variety of cell types. |
| RNA | abbreviation for ribonucleic acid, a class of single-stranded molecules involved in protein synthesis. |
| senescence | a state of being old, in which normal function is declining as death approaches. |
| trimerisation | the process of forming a substance that is composed of three molecules of a monomer. |
References
- The Journal Cell Death and Differentiation was started in January 1997. In terms of its ‘impact factor’ this is, as the editorial board claim, ‘the primary journal focused on the exciting field of programmed cell death and apoptosis’. The stated scope is ‘ … a journal devoted to the cell biology, molecular biology and biochemistry of cell death and differentiation, both in normal tissue regulation and in disease. To this end, Cell Death and Differentiation provides a unified forum for scientists as well as clinicians. It is committed to the rapid publication of high quality original papers that relate to these subjects, together with topical, usually solicited, reviews, meeting reports, editorial correspondence and occasional commentaries on controversial and scientifically informative issues’. Return to Text.
- Apoptosis: An International Journal on Programmed Cell Death was started in 1997. The stated aims and scope of this bimonthly periodical are: ‘Apoptosis is an international peer-reviewed journal, published bimonthly. The Journal is devoted to the rapid publication of innovative basic and clinically-oriented investigations into programmed cell death. It aims to stimulate both research on the basis of mechanisms of apoptosis and on its role in various human disease processes including: cancer, autoimmune disease, viral infection, AIDS, cardiovascular disease, neurodegenerative disorders, osteoporosis and ageing.’ Return to Text.
- Peter, M.E., Heufelder, A.E. and Hengartner, M.O., Advances in apoptosis research, Proc. Nat. Acad. Sci. USA 94:12736–12737, 1997. Return to Text.
- Kerr, J.F., Wylie, A.H. and Currie, A.R., Apoptosis: a basic biological phenomenon with wide-ranging implications in tissue kinetics, British J. Cancer 26(4):239–257, 1972. Return to Text.
- Wallach, D., Placing death under control, Nature 388:123–126, 1997. Return to Text.
- Behe, M.J., Rube Goldberg in the blood, Chapter 4; in: Darwin’s Black Box, The Free Press, Simon & Schuster Inc., New York, pp. 74–97, 1996. Return to Text.
- Walport, M., Complement; in: Roitt, I., Brostoff, J. and Male, D. (Eds), Immunology, Mosby, Times Mirror International Publishers Limited, London, pp. 13.1–13.17, 1996. Return to Text.
- Ellis, H. and Horvitz, H., Genetic control of programmed cell death in the nematode C. elegans, Cell 44:817–829, 1986. Return to Text.
- Hengartner, M., Ellis, R. and Horvitz, H., Caenorhabditis elegans gene ced-9 protects cells from programmed cell death, Nature 356:494–499, 1992. Return to Text.
- Edginton, S.M., Looking death in the eye: apoptosis and cancer research, Bio/Technology 11:787–792, 1993. Return to Text.
- Garfield, E. and Melino, G., The growth of the cell death field: an analysis from the ISI-Science citation index, Cell Death and Differentiation 4:352–361, 1997. Return to Text.
- Figures were obtained from online searches of the U.S. National Library of Medicine’s PubMed, <www.ncbi.nlm.nih.gov/PubMed/>. Return to Text.
- Bosanquet, A.G., Bell, P.B. and Rooney, N., Effect of interleukin-2 on CD95(Fas/APO-1) expression in fresh chronic lymphocytic leukaemia and mantle cell lymphoma cells: relationship to ex vivo chemoresponse, Anticancer Research 19:5329–5334, 1999. Return to Text.
- Cleveland, J.L. and Ihle, J.N., Contenders in FasL/TNF death signalling, Cell 81:479–482, 1995. Return to Text.
- Nagata, S. and Golstein, P., The Fas death factor. Science 267:1499–1456, 1995. Return to Text.
- Boldin, M.P., Varfolo, E.E., Pancer, Z., Mett, I.L., Camonis, J.H. and Wallach, D., A novel protein that interacts with the death domain of Fas/APO1 contains a sequence motif related to the death domain, J. Biological Chemistry 270:7795–7798, 1995. Return to Text.
- Chinnaiyan, A.M., O’Rourke, K., Tewari, M. and Dixit, V.M., FADD, a novel death domain-containing protein, interacts with the death domain of Fas and initiates apoptosis, Cell 81:505–512, 1995. Return to Text.
- Chinnaiyan, A.M., Tepper, C.G., Seldin, M.F., O’Rourke, K., Kischkel, F.C., Hellbardt, S., Krammer, P.H., Peter, M.E. and Dixit, V.M., FADD/MORT1 is a common mediator of CD95(Fas/APO-1) and tumor necrosis factor receptor-induced apoptosis, J. Biological Chemistry 271:4961–4965, 1996. Return to Text.
- Duan, H. and Dixit, V.M., RAIDD is a new ‘death’ adapter molecule, Nature 385:86–89, 1997. Return to Text.
- Alnemri, E.S., Livingston, D.J., Nicholson, D.W., Salvesen, G., Thornberry, N.A., Wong, W.W. and Yuan, J., Human ICE/CED-3 protease nomenclature, Cell 87:171, 1996. Return to Text.
- McGill, G. and Fisher, D.E., Apoptosis in tumorigenesis and cancer therapy, Frontiers in Bioscience, 2:d353–379, 1997. Return to Text.
- Bossy-Wetzel, E. and Green D.R., Apoptosis: checkpoint at the mitochondrial frontier, Mutation Research 434(3):243–251, 1999. Return to Text.
- Hancock, J.T., Desikan, R. and Neill S.J., Does the redox status of cytochrome C act as a fail-safe mechanism in the regulation of programmed cell death? Free Radic Biol Med 31(5):697–703, 2001. Return to Text.
- Mignotte, B. and Vayssiere, J.L., Mitochondria and apoptosis, European J. Biochemistry 252(1):1–15, 1998. Return to Text.
- Korsmeyer, S.J., Regulators of cell death, Trends in Genetics 11:101–105, 1995. Return to Text.
- Rao, L. and White, E., Bcl-2 and ICE family of apoptotic regulators: making a connection, Current Opinion in Genetic Development 7(1):52–58, 1997. Return to Text.
- The p53 tumour suppressor gene is necessary for many forms of apoptosis, induced only when DNA-strand breakage occurs and arresting the cell cycle in the G1 phase. As an essential cell cycle regulator it has been called the ‘guardian of the genome’. Aberrant p53 (i.e. it has sustained a somatic mutation) results in cells that fail to be driven into apoptosis; one cause of neoplastic cells that result in cancers developing. Two recent reviews on the relationship between p53 and apoptosis are: Wynford-Thomas, D., p53: guardian of cellular senescence, J. Pathology 180(2):118–121, 1996; Itahana, K., Dimri, G. and Campisi, J., Regulation of cellular senescence by p53, European J. Biochemistry 268(10):2784–2791, 2001. For a useful discussion of cell cycle control from a creationist perspective, see: Bergman, J., Cell cycle control and Paley’s watch, Journal of Creation 11(1):82–92, 1997. Return to Text.
- Rb, (retinoblastoma protein) and other functionally related proteins, are critical regulators of cell proliferation and apoptosis. Rb was first identified in relation to a rare pediatric eye tumour (an hereditary form of retinoblastoma). It is important in cancer research, as tumour cells that are resistant to chemotherapy typically have inactivated Rb pathways. A useful review is: Ping Dou, Q., Putative roles of retinoblastoma protein in apoptosis, Apoptosis 2:5–18, 1997. Return to Text.
- Perforin (also called PFP, for ‘pore-forming protein’) is an important mediator of the cytotoxic activity of ‘killer lymphocytes’ in the body’s immune system. See: Stepp, S.E., Matthew, P.A., Bennett, M., de Saint Basile, G. and Kumar, V., Perforin: more than just an effector molecule, Immunology Today 21(6):254–256, 2000. Return to Text.
- Granzymes are a family of serine proteases that function synergistically with perforins (see previous reference) to bring about the apoptosis of cells targeted by the immune system’s cytotoxic lymphocytes (natural killer [NK] cells and cytotoxic T cells [CTLs]). Good reviews of this topic are: Smyth, M.J. and Trapani, J.A., Granzymes: exogenous proteinases that induce target cell apoptosis, Immunology Today 16(4):202–206, 1995; Greenberg, A.H., Granzyme B-induced apoptosis, Advances in Experimental and Medical Biology, 406:219–228, 1996. Return to Text.
- Ceramide mediates an intracellular messenger that has key roles in cell-growth suppression, cell differentiation, cell-cycle arrest and in mediating apoptosis. See: Perry, D.K., Ceramide and apoptosis, Biochemical Society Transactions 27(4):399–404, 1999. Return to Text.
- Morris, H., The Genesis Record: A scientific & devotional commentary on the book of beginnings, Baker Book House, Grand Rapids, Michigan, pp. 69–71, 1976. Return to Text.
- Ham, K., Demolishing ‘straw men’, Creation 19(4):13–15, 1997; especially p. 14. Return to Text.
- Sarfati, J., Genesis questioned by billions of years beliefs, Journal of Creation 13(2):22–30, 1999; especially p. 29. Return to Text.
- Batten, D. (Ed.), Ham, K., Sarfati, J. and Wieland, C., The Answers Book. Answers in Genesis, Triune Press, Brisbane, pp. 32 and 92, 1999. Return to Text.
- Since this genetic programme is translated by the cellular machinery to effect precise cellular changes that accomplish the purpose of cell removal, the encoded information clearly fulfils the criteria of the fifth (highest) level of Information Theory. This is further evidence for a Supreme Source of Intelligence, as demonstrated in Werner Gitt’s seminal book: Gitt, W., In the Beginning was Information, Christliche Literatur-Verbreitung e.V., Bielefeld, Germany, 1997. Return to Text.
- Searle, J., Kerr, J.F. and Bishop, C.J., Necrosis and apoptosis: distinct modes of cell death with fundamentally different significance, Pathology Annual 17:229–259, 1982. Return to Text.
- Buja, L.M., Eigenbrodt, M.L. and Eigenbrodt, E.H., Apoptosis and necrosis. Basic types and mechanisms of cell death, Archives of Pathological and Laboratory Medicine 117(12):1208–1214, 1993. Return to Text.
- Shi, Y.F., Szalay, M.G., Paskar, L., Sahai, B.M., Boyer, M., Singh, B. and Green, D.R., Activation-induced cell death in T cell hybridomas is due to apoptosis. Morphological aspects and DNA fragmentation, J. Immunology 144(9):3326–3333, 1990. Return to Text.
- Trump, B.F. and Berezesky, I.K., The role of cytosolic Ca2+ in cell injury, necrosis and apoptosis, Current Opinion in Cell Biology 4(2):227–232, 1992. Return to Text.
- Tomei, L.D., Kiecolt-Glaser, J.K., Kennedy, S. and Glaser, R., Psychological stress and phorbol ester inhibition of radiation-induced apoptosis in human peripheral blood leukocytes, Psychiatry Research 33(1):59–71, 1990. Return to Text.
- Wylie, A.H., Glucocorticoid-induced thymocyte apoptosis is associated with endogenous endonuclease activation, Nature 284(5756):555–556, 1980. Return to Text.
- Gregoire, V., Van, N.T., Stephens, L.C., Brock, W.A., Milas, L., Plunkett, W. and Hittelman, W.N., The role of fludarabine-induced apoptosis and cell cycle synchronization in enhanced murine tumor radiation response in vivo, Cancer Research 54(23):6201–6209, 1994. Return to Text.
- Arends, M.J., Morris, R.G. and Wylie, A.H., Apoptosis. The role of the endonuclease, American J. Pathology 136(3):593–608, 1990. Return to Text.
- Barry, M.A. and Eastman, A., Identification of deoxyribonuclease II as an endonuclease involved in apoptosis, Archives of Biochemistry and Biophysics 300(1):440–450, 1993. Return to Text.
- Williams, G.T. and Smith, C.A., Molecular regulation of apoptosis: genetic controls on cell death, Cell 74(5):777–779, 1993. Return to Text.
- Tanaka, R., Takeda, N., Okada, K. and Ueki, K., Computerized tomography of coagulation necrosis of the brain and brain tumors, Surgical Neurology 11(1):9–12, 1979. Return to Text.
- Duarte-Karim, M., Ruysschaert, J.M. and Hildebrand, J., Affinity of adriamycin to phospholipids. A possible explanation for cardiac mitochondrial lesions, Biochemical and Biophysical Research Communications 71(2):658–663, 1976. Return to Text.
- Shiratsuchi, A., Umeda, M., Ohba, Y. and Nakanishi, Y., Recognition of phospatidylserine on the surface of apoptotic spermatogenic cells and subsequent phagocytosis by sertoli cells of the rat, J. Biological Chemistry 272(4):2354–2358, 1997. Return to Text.
- Lockshin, R.A., The early modern period in cell death, Cell Death and Differentiation 4:347–351, 1997. Return to Text.
- Mori, C., Nakamura, N., Kimura, S., Irie, H., Takigawa, T. and Shiota, K., Programmed cell death in the interdigital tissue of the fetal mouse limb is apoptosis with DNA fragmentation, Anatomical Records 242(1):103–110, 1995. Return to Text.
- Thomazy, V.A. and Davies, P.J., Expression of tissue transglutaminase in the developing chicken limb is associated both with apoptosis and endochrondral ossification, Cell Death and Differentiation 6(2):146–154, 1999. Return to Text.
- Cohen, J.J., Programmed cell death and apoptosis in lymphocyte development and function, Chest 103(2Suppl):99S–101S, 1993. Return to Text.
- D’Adamio, L., Awad, K.M. and Reinherz, E.L., Thymic and peripheral apoptosis of antigen-specific T cells might cooperate in establishing self tolerance, European J. Immunology 23(3):747–753, 1993. Return to Text.
- Neiman, P.E., Thomas, S.J. and Loring, G., Induction of apoptosis during normal and neoplastic B-cell development in the bursa of Fabricius, Proc. Nat. Acad. Sci. USA 88(13):5857–5861, 1991. Return to Text.
- Lu, L. and Osmond, D.G., Regulation of cell survival during B lymphopoiesis in mouse bone marrow: enhanced pre-B-cell apoptosis in CSF-1-deficient op/op mutant mice, Experimental Hematology 29(5):596–601, 2001. Return to Text.
- Griffith, T.S., Brunner, T., Fletcher, S.M., Green, D.R. and Ferguson, T.A., Fas/Fas-ligand-induced apoptosis as a mechanism of immune privilege, Science 270(5239):1189–1192, 1995. Return to Text.
- Kasahara, Y., Iwai, K., Yachie, A., Ohta, K., Konno, A., Seki, H., Miyawaki, T. and Taniguchi, N., Involvement of reactive oxygen intermediates in spontaneous and CD95(Fas/APO-1)-mediated apoptosis of neutrophils, Blood 89(5):1748–1753, 1997. Return to Text.
- Dunkel, L., Hirvonen, V. and Erkkila, K., Clinical aspects of male germ cell apoptosis during testis development and spermatogenesis, Cell Death and Differentiation 4:171–179, 1997. Return to Text.
- Rockett, J.C., Mapp, F.L., Garges, J.B., Luft, J.C., Mori, C. and Dix, D.J., Effects of hyperthermia on spermatogenesis, apoptosis, gene expression, and fertility in adult male mice, Biology of Reproduction 65(1):229–239, 2001. Return to Text.
- Tilly, J.L., Tilly, K.I. and Perez, G.I., The genes of cell death and cellular susceptibility to apoptosis in the ovary: a hypothesis, Cell Death and Differentiation 4:180–187, 1997. Return to Text.
- Andres, A.C. and Strange, R., Apoptosis in the estrous and menstrual cycles, J. Mammary Gland Biology and Neoplasia 4(2):221–228, 1999. Return to Text.
- Shikone, T., Kokawa, K., Yamoto, M. and Nakano, R., Apoptosis of human ovary and uterine endometrium during the menstrual cycle, Hormone Research 48(Suppl 3):27–34, 1997. Return to Text.
- Wilde, C.J., Knight, C.H. and Flint, D.J., Control of milk secretion and apoptosis during mammary involution, J. Mammary Gland Biology and Neoplasia 4(2):129–136, 1999. Return to Text.
- Hakansson, A., Zhivotovsky, B., Orrenius, S., Sabharwal, H. and Svanborg, C., Apoptosis induced by a human milk protein, Proc. Nat. Acad. Sci. USA 92:8064–8068, 1995. Return to Text.
- Bell, P.B., Unpublished experiments, December 1995. I tested serial dilutions of several breast-milk specimens (from healthy women) against both leukaemic and normal lymphocytes. For two of the milk samples, a concentration as low as 0.625% breast milk (diluted in phosphate buffered saline) was sufficient to obliterate the majority of malignant lymphocytes (from two patients with chronic lymphocytic leukaemia) in a 4-day culture (compared to untreated leukaemic cell controls). There was less cytotoxic activity against normal lymphocytes from two non-leukaemic specimens (peripheral blood specimens from two of my colleagues). Return to Text.
- Berger, A., Milk protein may help beat cancer, New Scientist 147(1992):15, 1995. Return to Text.
- Damjanovski, S., Amano, T., Li, Q., Ueda, S., Shi, Y.B. and Ishizuya-Oka, A., Role of ECM remodelling in thyroid hormone-dependent apoptosis during anuran metamorphosis, Annals of the New York Academy of Science 926:180–191, 2000. Return to Text.
- Lockshin, R.A., Zakeri, Z. and Tilly, J.L. (Eds), When Cells Die: A Comprehensive Evaluation of Apoptosis and Programmed Cell Death, Wiley-Liss, New York, 1998. Return to Text.
- Tomei, L.D. and Umansky, S.R., Aging and apoptosis control, Neurol. Clin. 16(3):735–745, 1998. Return to Text.
- Michie, C.A. and McClean, A., Lymphocyte longevity: a coincidence of approaches, International J. Radiation Biology 66(2):229, 1994. Return to Text.
- Bergman, J., The life clock and Paley’s watch: The telomeres, CRSQ 37(3):176–183, 2000. Return to Text.
- Bergman, Ref. 72, p. 181. Return to Text.
- Behe, M.J., Darwin’s Black Box, The Free Press, Simon & Schuster Inc., New York, 1996. Return to Text.
- McGill, G. and Fisher, D.E., Apoptosis in tumorigenesis and cancer therapy, Frontiers in Bioscience 2:d353–379, 1997. Return to Text.
- Lowe, S. and Ruley, E., p-53-dependent apoptosis in tumor progression and in cancer therapy; in: Holbrook, N., Martin, G. and Lockshin, R. A. (Eds), Cellular Aging and Cell Death, Wiley-Liss, New York, pp. 209–234, 1996. Return to Text.
- Tsujimoto, Y., Cossman, J., Jaffe, E. and Croce, E.M., Involvement of the bcl-2 gene in human follicular lymphoma, Science 228:1440–1443, 1985. Return to Text.
- Delia, D., Aiello, A., Soligo, D., Fontanella, E., Melani, C., Pezzella, F., Pierotti, M.A. and Della Porta, G., bcl-2 proto-oncogene expression in normal and neoplastic human myeloid cells, Blood 79:1291–1298, 1992. Return to Text.
- Hsu, B., Marin, M.C., Brisbay, S., McConnell, K. and McDonnell, T.J., Expression of bcl-2 gene confers multidrug resistance to chemotherapy-induced cell death, The Cancer Bulletin 46:125–129, 1994. Return to Text.
- Mitra, D., Steiner, M., Lynch, D.H., Staiano-Coico, L. and Laurence, J., HIV-1 upregulates Fas ligand expression in CD4+ T cells in vitro and in vivo: association with Fas-mediated apoptosis and modulation by aurintricarboxylic acid, Immunology 87(4):581–585, 1996. Return to Text.
- Plymale, D.R., Tang, D.S., Comardelle, A.M., Fermin, C.D., Lewis, D.E. and Garry, R.F., Both necrosis and apoptosis contribute to HIV-1-induced killing of CD4 cells, AIDS 13(14):1827–1839, 1999. Return to Text.
- Jaworowski, A. and Crowe, S.M., Does HIV cause depletion of CD4+ T cells in vivo by the induction of apoptosis? Immunological Cell Biology 77(1):90–98, 1999. Return to Text.
- Wolozin, Z., Neuronal loss in aging and disease; in: Holbrook, N., Martin, G. and Lockshin, R.A. (Eds), Cellular aging and cell death, Wiley-Liss, New York, p. 289–300, 1996. Return to Text.
- Roth, K.A., Caspases, apoptosis, and Alzheimer’s disease: Causation, correlation, and confusion, J. Neuropathology and Experimental Neurology 60(9):829–838, 2001. Return to Text.
- Raina, A.K., Hochman, A., Xiongwei, Z., Rottkamp, C.A., Nunomura, A., Siedlak, S.L., Boux, H., Castellani, R.J., Perry, G. and Smith, M.A., Abortive apoptosis in Alzheimer’s disease, Acta Neuropathalogica 101(4):305–310, 2001. Return to Text.
- Yatsugi, N., Tsukazaki, T., Osaki, M., Koji, T., Yamashita, S. and Shindo, H., Apoptosis of articular chondrocytes in rheumatoid arthritis and osteoarthritis: correlation of apoptosis with degree of cartilage destruction and expression of apoptosis-related proteins of p53 and c-myc, J. Orthopaedic Science 5(2):150–156, 2000. Return to Text.
- Chou, C.T., Yang, J.S. and Lee, M.R., Apoptosis in rheumatoid arthritis—expression of Fas, Fas-L, p53 and Bcl-2 in rheumatoid synovial tissues, J. Pathology 193(1):110–116, 2001. Return to Text.
- Mountz, J.D., Hsu, H., Matsuki, Y. and Zhang, H., Apoptosis and rheumatoid arthritis: past, present and future directions, Current Rheumatological Reports 3(1):70–78, 2001. Return to Text.
- Li, K., Li, Y., Shelton, J.M., Richardson, J.A., Spencer, E., Chen, Z.J., Wang, X. and Williams, R.S., Cytochrome c deficiency causes embryonic lethality and attenuates stress-induced apoptosis, Cell 101(4):389–399, 2000. Return to Text.
- Rosenfeld, M.E., Prichard, L., Shiojiri, N. and Fausto, N., Prevention of hepatic apoptosis and embryonic lethality in RelA/TNFR-1 double knockout mice, American J. Pathology 156(3):997–1007, 2000. Return to Text.
- Wilson, S.E., Stimulus specific and cell type-specific cascades: emerging principles relating to control of apoptosis in the eye, Experimental Eye Research 69(3):255–266, 1999. Return to Text.
- Lockshin et al., Ref. 69, from the preface. Return to Text.
- Demick, D., Cancer and the curse, Back to Genesis (ICR) 145, January 2001. Return to Text.
- Bosanquet, A.G. and Bell, P.B., Novel ex vivo analysis of nonclassical, pleiotropic drug resistance and collateral sensitivity induced by therapy provides a rationale for treatment strategies in chronic lymphocytic leukaemia, Blood 87(5):1962–1971, 1996. Return to Text.
- Bosanquet, A.G. and Bell, P.B., Enhanced ex vivo drug sensitivity testing of chronic lymphocytic leukaemia using refined DiSC assay methodology, Leukemia Research 20(2):143–153, 1996. Return to Text.
- Bell, P.B., Rooney, N. and Bosanquet, A.G., CD79a detected by ZL7.4 separates chronic lymphocytic leukaemia from mantle cell lymphoma in the leukemic phase, Cytometry (Communications in Clinical Cytometry) 38:102–105, 1999. Return to Text.
- Bosanquet et al., Ref. 13. In contrast to previous reports, which used supra-pharmacological interleukin-2 (IL-2) concentrations (>1000 u/ml), we used a much lower concentration—sufficient for T-cell activation—to mimic in vivo plasma concentrations. We found that IL-2 had a variable effect on Fas-levels in chronic lymphocytic leukaemia (CLL) cells—either significant up-regulation or no change—but no effect on Fas-levels in mantle cell lymphoma (MCL) cells. Contrary to expectations, where Fas-induction occurred, this was associated with marked ex vivo multi-drug resistance to cytotoxic drugs, indicating that Fas-induction might be a negative prognostic indicator in a subset of CLL patients. Return to Text.
- White, E., Overview of apoptosis, in: Apoptosis: A Complete Catalog and Scientific Reference Guide, Oncogene Research Products, Calbiochem, 1996. Return to Text.
- Anon, News item for 30 January 1997 from the home page of The New York Area Cell Death Society; <http://www.celldeath-apoptosis.org> accessed 6 February 1997. Return to Text.
- Apoptosis (albeit in response to non-physiological triggers, such as irradiation) has been described in several unicellular eukaryotes. The evolutionist therefore has to explain how it evolved before the invention of multicellularity. The intractable dilemma for the evolutionist is this: how would a functional apoptotic response in a unicellular organism ever have been selected for? What selection advantage is there in eliminating oneself? Return to Text.
- Bell, P.B., The non-evolution of apoptosis (in preparation). This deals with the alleged evolutionary conservation of apoptosis, from the earliest eukaryotic cells. Return to Text.
- Burgess, S., Extreme diversity; in: Hallmarks of Design: Evidence of Design in the Natural World, Day One Publications, Epsom, Great Britain, pp. 101–118, 2000. Return to Text.
- Guide to Cell Proliferation and Apotosis Methods, Hoehringer Mannheim, Mannheim, p. 30, 1997. Return to Text.


Readers’ comments
Comments are automatically closed 14 days after publication.