

Journal of Creation 16(2):13–15, August 2002
Browse our latest digital issue Subscribe
C4 photosynthesis—evolution or design?

Life depends on photosynthesis, where plants take carbon dioxide from the atmosphere and ‘fix’ it into high-energy sugars using light as the energy source. Two basic forms of photosynthesis have been discovered. In one, the first compound made from CO2 is a three-carbon compound, so this is called C3 photosynthesis. In the other, the first compound is a four-carbon compound, so it is called C4 photosynthesis.1 Most plants are C3; about 15% of species have the C4 system. Examples of C3 plants include wheat, rice, potatoes and cabbage. C4 plants include maize, sugar cane, sorghum and succulents—mainly tropical/arid environment species.
C4 and C3 plants differ in their leaf anatomy and where photosynthesis occurs. C3 plants have chloroplasts throughout the internal (‘mesophyll’) leaf cells, and there are air spaces around the cells to allow ready diffusion of CO2 into them. In C4 plants, the photosynthetic cells cluster around the vascular bundles (leaf veins) and there are no air spaces around the photosynthetic cells. The photosynthetic cells are called bundle sheath cells because they form a tight sheath around the vascular bundles.
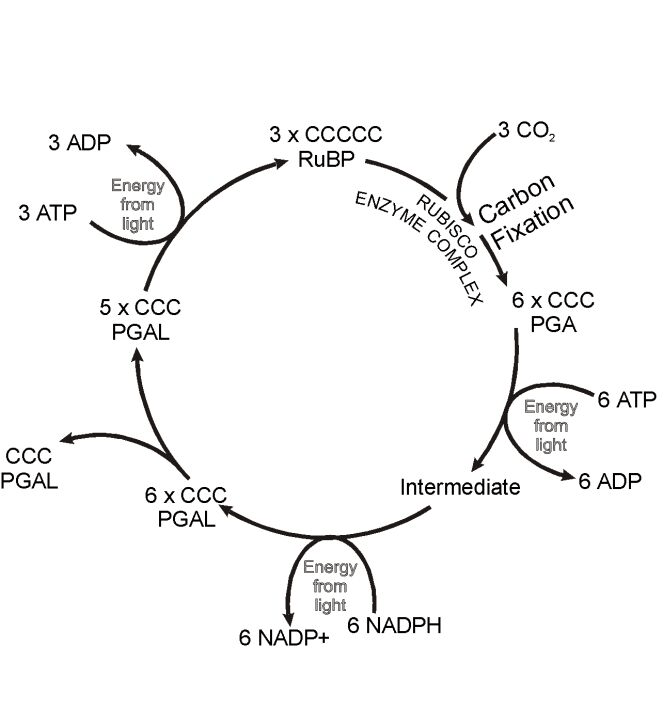
C3 and C4 plants share the same light-harvesting systems, as well as the same enzyme cycle for incorporating the carbon into sugars—the Calvin-Benson cycle. The first enzyme in this cycle, nicknamed ‘Rubisco’, makes up 25% of the protein in leaves, which makes it the most abundant protein on Earth. Rubisco takes CO2 and adds it to a 5-carbon sugar, making two
C4 plants have extra enzymes operating in the leaf. These incorporate the CO2 (actually bicarbonate, HCO3–) into a
C3 and C4 plants also differ in that C3 plants exhibit ‘photorespiration’, where they lose some of the CO2 fixed into
This seems to be a safety mechanism to avoid damage to the photosynthesis system at low CO2 levels. If there is inadequate CO2 to fix the energy harvested by the chlorophyll system, then oxygen radicals form and these damage the light harvesting system. Photorespiration maintains a supply of ADP and NADP to accept the energy generated by the light-harvesting system.
C4 plants concentrate their CO2, thus suppressing photorespiration. Also, since the supply of CO2 is maintained, even at low concentrations, there is always a sink for the energy from the light harvesting, so damage to the photosystems is avoided. So there is no need for photorespiration.
Why two methods of fixing CO2?
Why do C3 plants tend to be temperate in their adaptation and C4 plants tropical/arid? The rate of photorespiration rises rapidly with temperature, so it becomes a much more serious problem, in terms of its ‘inefficiency’ (loss of fixed carbon), in the tropics. On the other hand, the C4 system has energy costs: each CO2 fixed into malate needs one NADPH and one ATP for the complete cycle. So the relative advantages seem to be due to the trade-off between photorespiration in C3 plants and the extra costs of carbon fixation in C4 plants. With increasing temperatures, the cost of photorespiration becomes greater than the extra cost of the C4 system, which is met by the increased sunlight energy anyway, so the latter prevails.
C4 plants also do well in arid environments. In this situation the plant closes its stomata (leaf pores) to conserve water. This also reduces the amount of CO2 entering the leaf and raises the leaf temperature. The enzyme that fixes CO2 in C4 plants has a much greater affinity for CO2 than Rubisco, which does the job alone in C3 plants. So C4 plants are still able to supply plenty of CO2 to the Rubisco in the photosynthetic cells, whereas a C3 plant would have trouble.2
The origins of the C4 system
Some 8,000 to 10,000 species of plants in 18 families, including both monocots (which includes grasses) and dicots (roughly, ‘broad-leaved’ plants), have the C4 system. C4 metabolism has even been found in a single-celled marine diatom.3
Many flowering plant families have both C3 and C4 species. Some species are intermediate, showing both C3 and C4 characteristics. In the Atriplex genus, some species are C3, while others are C4, and C3 and C4 species have been hybridized.4 Wood and Cavanaugh have reviewed the genus Flaveria, which has species of C3, C4 and intermediate type, many of which hybridize.5
The distribution of C4 species does not form any pattern that could relate to any reasonable evolutionary phylogeny. Consequently, evolutionists have proposed that C4 photosynthesis has arisen independently at least 30 times—a classic case of ‘polyphyletic evolution’.
However, C4 chemistry involves several complex enzyme systems and the chemistry is remarkably consistent across the spectrum (there are three types of enzyme used to release the CO2 from the organic acid that transports the CO2, otherwise the chemistry is similar).
To believe that C4 chemistry arose once by natural processes would require super ‘faith’ for the evolutionist. But to propose that such a system with its new complex coded genetic information arose separately some 30 times by mutations and natural selection, and that these processes arrived at essentially the same solution, stretches credulity to breaking point. This would be an extreme example of ‘convergent evolution’—even more than the supposed polyphyletic origin of the eye in general and the compound eye in particular.6
Some species that exhibit both C3 and C4 forms are even able to switch from one to the other during development. This suggests that maybe the C4 chemistry is latent in C3 plants, or is suppressed by some means. In the marine diatom mentioned above, C4 metabolism seems to be facultative.3
Wood and Cavanaugh5 concluded from their baraminological study of Flaveria that the C4 photosynthetic pathway arose from plants that were originally C3, and this probably happened post-Flood. These authors propose that the genetic information for C4-mode photosynthesis was present in the original created kinds, but latent and has become activated since.
Surprise: C3 plants have the C4 system!
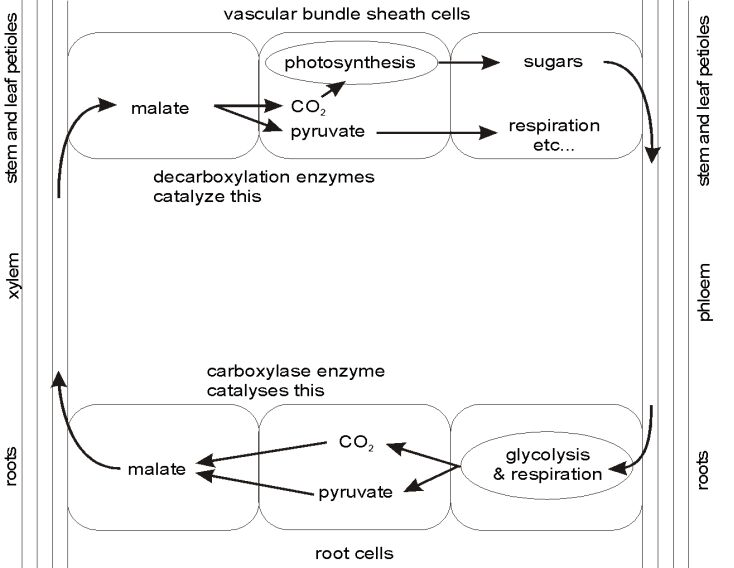
Now Hibberd and Quick have shown that tobacco and celery, two classical C3 plants, contain virtually all the C4 characteristics, not in their leaves, but in their roots, stems and petioles.7 They showed that CO2 respired in the roots is fixed into malate by the same enzyme that fixes CO2 in the leaves of C4 plants. The malate moves in the xylem stream up the plant where it transfers into bundle sheath cells surrounding the vascular bundles in the stems and petioles. Here all three decarboxylation (CO2-releasing) enzymes identified in the three sub-types of the C4 system are present in elevated levels. They release the CO2 so that Rubisco can use it in the Calvin cycle. The chemistry is apparently identical to the C4 system. These plants differ from C4 plants only in the site of synthesis of the malate (roots in C3 plants versus leaf mesophyll cells in C4 plants) and its transfer to the bundle sheath cells. Even the anatomy of the bundle sheath cells in the stems and petioles is similar.
This makes for a very efficient system for retrieving respired carbon from the roots. Indeed, CO2 may even enter the roots from the soil, where the level of CO2 is usually quite high due to the activity of heterotrophic micro-organisms. This would reduce the CO2 concentration in the soil, which would be beneficial to the aerobic organisms living there. What wonderful design for an efficient ecology!
Hibberd and Quick point out that since so much of the C4 system is already present in the C3 plants, ‘fewer modifications are needed for C4 photosynthesis to evolve’. Indeed, are we talking about the origin of new complex, coded genetic information at all, or are we looking at adaptation based on existing genetic information—as proposed by the creationists Wood and Cavanaugh?
It now seems that the genes for C4 enzymes and anatomy are selectively expressed in the roots, stems and petioles of C3 plants, but are suppressed in the leaves. C4 plants differ in having these genes expressed in the leaves as well. If the suppression in the leaves of C3 plants were due to the synthesis of proteins that interact with promoter sequences, for example, it may even be possible to see mutations in the genes for these proteins that result in the expression of C3–C4 or C4 photosynthesis. Or there might be some designed means of switching on this adaptation genetically so that it is inherited once switched on—something like Wood’s Altruistic Genetic Elements (AGEs)?5
These developments underline just how cleverly the original plants were created—with built-in latent capacity for adaptation to a wide range of environments. It will be interesting to see the details fleshed out.
References
- The basic details of C4 photosynthesis were elucidated by Australian scientists in the 1960s. See Hatch, M.D. and Slack, C.R., Photosynthesis by sugarcane leaves, Biochem. J. 101:103–111, 1966. Return to Text.
- A variation on the C4 theme is seen in CAM (crassulacean acid metabolism) plants. Typically succulent desert plants, they open their stomata at night to fix CO2, storing the fixed form in vacuoles (reservoirs within cells), then release the CO2 for photosynthesis during the day when the stomata shut. In this manner they conserve water very efficiently. Return to Text.
- Reinfelder, J.R., Kraepiel, A.M.L. and More, F.M.M., Unicellular C4 photosynthesis in a marine diatom, Nature 407:996–999, 2000. Return to Text.
- Sengbusch, P. v., Influence of different parameters on the efficiency of the CO2-uptake—C3-and a C4-plant, <www.biologie. uni-hamburg.de/b-online/e24/8.htm>, accessed 15 March 2002. Return to Text.
- Wood, T.C. and Cavanaugh, D.P., A baraminological analysis of subtribe Flaverinae (Asteraceae: Helenieae) and the origin of biological complexity, Origins (GRI) 52:7–27, 2001. Return to Text.
- Oakley, T.H. and Cunningham, C.W., Molecular phylogenetic evidence for the independent evolutionary origin of an arthropod compound eye, Proc. Nat. Acad. Sci. USA 99(3):1426–1430, 2002. Their abstract says, ‘These results illustrate exactly why arthropod compound eye evolution has remained controversial, because one of two seemingly very unlikely evolutionary histories must be true. Either compound eyes with detailed similarities evolved multiple times in different arthropod groups or compound eyes have been lost in a seemingly inordinate number of arthropod lineages.’ Return to Text.
- Hibberd, J.M. and Quick, W.P., Characteristics of C4 photosynthesis in stems and petioles of C3 flowering plants. Nature 415:451–453, 2002. Return to Text.
Readers’ comments
Comments are automatically closed 14 days after publication.