

Journal of Creation 15(3):92–99, December 2001
Browse our latest digital issue Subscribe
Dawkins’ eye revisited
This article is written in response to comments Richard Dawkins has made concerning the human eye. Besides disparaging the organisation of the retina, he also claims that the eye could have developed gradually by small increments. The anatomy and physiology of the cornea, the eyelids and the tears illustrate how his reasoning is fallacious because of the principles of irreducible complexity and of genetic information gain.
In his widely acclaimed book The Blind Watchmaker,1 Dawkins sets out to convince his readers that the staggering complexity and diversity of biological life that surrounds us in nature came into being by the gradual accumulation of very small advances over many millions of years through mutations coupled with natural selection. In an attempt to counter William Paley’s arguments for design in the human eye, he devotes much attention to this organ as a prime example (as he supposes) of only apparent design. And so he denigrates the Creator (p. 93) for bungling (in his view) the design of the retina with its ‘back-to-front’ wiring which he labels as a ‘revealing flaw’. His remarks with regard to the retina have been rebutted in previous publications of Creation Ministries International.2–4
Earlier in his same book, Dawkins quotes (p. 80) from Francis Hitching’s book called The Neck of the Giraffe or Where Darwin Went Wrong.5 Hitchings makes the valid point about our visual system that for it to function, many complex components must operate simultaneously and that failure of one component can disable it. How, therefore, could any of its essential components come into existence without the others for any of them to be of benefit to the creature? Dawkins then comments as follows:
‘This remarkable argument is very frequently made, presumably because people want to believe its conclusion’ (p. 80).
Click here for larger view
However true this may be, Dawkins here seems to be unaware of his own prejudice: he likewise wants to believe macroevolution and to destroy creationism with a zeal and fervour to which his many adulators readily testify. Significantly, Dawkins is unable to refute the thrust of Hitching’s argument—variously termed by others as ‘correlated adaptation’ (Spetner6), ‘irreducible complexity’ (Behe7) or ‘irreducible mechanism’ (Burgess8). And so he immediately seizes on an unfortunate exaggeration or error made by Hitchings in order to distract attention from his own impotence. He uses the same technique (pp. 86–87) with Hitching’s discussion of the bombardier beetle and human lungs.
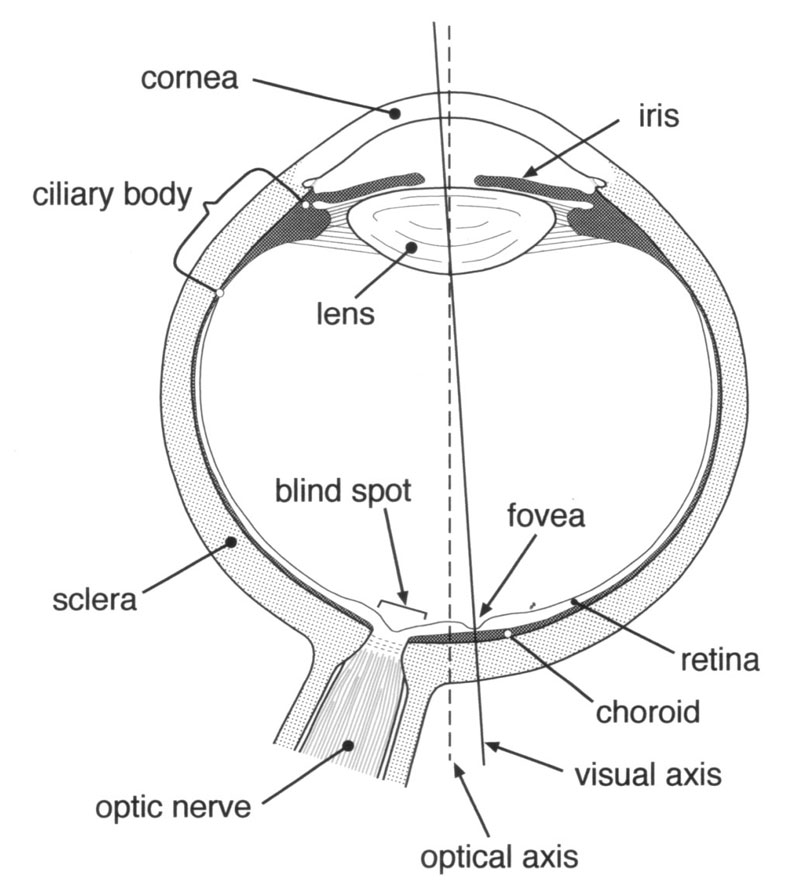
Returning to the eye (Figure 1), Hitchings could have avoided this counter-attack by comparing partial failure of a component with complete failure. Partial failure causes a greater or lesser degree of disablement of the system, depending on the particular component and on the nature and severity of its malfunction. On the other hand, complete failure or absence of one component in the system would disable the system completely. For example, disorders of the optic nerve reduce vision by various degrees but severing the nerve renders the eye completely blind.
The ‘beginnings of vision’
In support of the notion that the eye could have evolved in small increments, Dawkins argues that 5% or 1% of normal vision is better than none at all and so by a process of small steps the eye’s performance could improve gradually over many generations.
‘Vision that is 5 per cent as good as yours or mine is very much worth having in comparison with no vision at all. So is 1 per cent vision better than total blindness. And 6 percent is better than 5, 7 better than 6, and so on up the gradual continuous series.’
This simplistic logic glosses over at least two major considerations. First, as indicated above, it ignores the principle of irreducible complexity. Let us imagine, for the sake of argument, some lowly creature for whom the avoidance of light would give it some supposed evolutionary advantage to evade a predator; and that it had acquired by evolution a single photoreceptor cell. Such an acquisition would, of itself, be a major achievement in that it is so specialized. But for that receptor to be of any value to the creature, a neural or humoral mechanism for communication and coordination within its body would have to be in place at the same time. All this immensely reduces the possibility of such a system ever evolving, not to mention the insuperable biochemical obstacles to the first appearance of a single cell.9–11 Furthermore, any predator of such a creature would need a sophisticated visual or other sensory system to detect the presence of the prey which then begs the question as to how the predator evolved! If there were no predator then the potential prey would not need the protection such a primitive visual system might afford. The visual system would then be eliminated by natural selection as being unneeded!
Secondly, any improvement in an animal’s genetic endowment as part of the supposed evolutionary process requires the incorporation of more information in its genome. This, as a spontaneous phenomenon, i.e. without intelligent intervention, has yet to be observed or reproduced experimentally and so remains a supposition, not an established fact. Observed mutations have never generated new information but, rather, they represent corruption of the genetic data, i.e. there is a loss of information.12
A recent editorial article13 has reviewed research with a bearing on the question of whether vision reduced to only perception of light is of value to those so afflicted. Workers who train and assist the visually handicapped agree that even this extremely poor level of vision is useful for orientation and mobility. A single light source can guide and orientate the subject. Light is differentially reflected off various surfaces, e.g. more from concrete, painted walls or crosswalks than from darker areas such as grass etc; this can provide clues for the handicapped. However, nonetheless, the fact remains that the severely visually impaired are very vulnerable and dependent on community goodwill. They are unable to avoid obstacles in their path when relying solely on their vision without the other senses.
Furthermore, unless the handicapped person can at the same time perceive the direction of light rays (known as light projection) entering the eye, the light in practice will be virtually no help for the purpose of mobility. Accurate light projection requires a large number of functioning photoreceptors suitably disposed to receive an image, together with their connections to the brain;14 it represents a huge advance in complexity over bare perception of light with no projection, not only in the eye but more so in the brain.
The same can be said for each successive stage in the supposed evolutionary development of our visual system. This would require the simultaneous acquisition of new structures in the eye plus increases in the complexity and organisation of the brain; one without the other would confer no competitive advantage in natural selection. It would entail progressing from perception of light with projection to the vaguest perception of movement, to form sense (the ability to perceive shapes), to colour vision, to binocular depth perception etc. Each stage is an increase in complexity of several orders of magnitude involving both eye and brain, to say nothing of the six muscles controlling the position and movement of each eye.
For any degree of worthwhile form sense, the eye’s optical system must form an image on the retina and this requires for vertebrates a clear refracting medium through which the light must pass before reaching the retina. This is provided in the human eye by the lens and the cornea.
To illustrate the two principles mentioned above (irreducible complexity and genetic information gain) it is instructive to consider the cornea, the eyelids and the tears.
The cornea

Click here for larger view
The internal natural lens of the eye is a device for fine tuning the refraction of the eye’s optical system, i.e. to vary its focal length according to the distance of the object being viewed. Now, it is true that the human eye can usefully function up to a point without its lens as Dawkins suggests, particularly in those who are very shortsighted (myopic),15 for whom removal of the lens can be beneficial.16 But this is only because of the presence of another more powerful lens of fixed focus, namely the cornea (see figures 1 & 2). Approximately two thirds of the refraction of light rays entering the eye occurs at the interface between air and the thin film of tears covering the front surface of the cornea.17 Thus because of the cornea the eye can still form an image on the retina, albeit a blurred one, in the absence of the natural lens.18 The refraction that normally occurs at the air/tear interface explains why, when the human eye is open underwater, the optical imagery is extremely blurred; the index of refraction of water is quite similar to that of the cornea and most of the optical power of the anterior corneal surface is lost.19 If the cornea with its covering of tears is to act as an optical lens it has to be transparent to light. As a transparent object the cornea is remarkable for its efficiency in this respect in that normally it transmits more than 90% of incident light.20 The cornea is part of the eyeball’s tough fibrous outermost tunic of which it forms the anterior one sixth in section, the other five sixths being the white opaque sclera (’the white of the eye’). The predominant material of this tunic is the fibrous protein collagen which is found throughout the body and is the main constituent e.g. of tendons.
Several factors are together responsible for the cornea’s high degree of transparency in contrast with the opaqueness of the sclera, and they reveal an extraordinary degree of organisation and precision. It is obvious that the absence of blood vessels and pigment in the cornea and the paucity of cells in its stroma (see below) are prerequisites for its transparency. The external surface membrane, the epithelium, is about 50 µm thick and consists of 5–6 layers of cells which are constantly being replaced from the cornea’s periphery. The epithelium is of homogeneous refractive index. When covered by a tear film it presents a perfectly smooth, almost spherical convex surface that does not scatter entering light. The corneal surface is the most specialized 123 mm2 of the body’s surface.21 Disease processes affecting the epithelium may cause unevenness of its surface which scatters incident light; they may also disturb its transmission and refraction of light, all of which can markedly reduce vision. Besides its optical properties, the superficial cells of the epithelium have tight junctions between them so that the epithelium is semipermeable; it functions as a barrier with a higher electrical resistance than that of the layers behind it. Important for the physiology of the deeper layers, it also controls the absorption of fluid from tears, medications etc, besides resisting invasion by microorganisms.
The basal cells of the epithelium are firmly adherent to their common basement membrane by means of hemidesmosomes (hemi + Greek desmos, a band + Greek soma, body). The basement membrane in turn is anchored by fibrils22 which reach into and, like Velcro, tangle with and grip the fibrils of the underlying transparent membrane (Bowman’s—see below). The epithelium is constantly being replaced, the basal cells giving rise to the superficial cells; the basal cells also are constantly being produced by stem cells23,24 located at the junctional zone between sclera and cornea, known as the limbus. The basal cells migrate over the cornea centripetally, marching in columns,25 from the limbus; and the superficial cells are shed slightly below the cornea’s central area. Animal experiments have revealed that when epithelium is lost with an injury, a coordinated response is triggered to close the defect: the normal movement of epithelium is accelerated with the cells flattening and the basal cells in the vicinity of the wound losing their hemidesmosomes within two hours.26
Bowman’s membrane to which the epithelium is attached is about 12 µm thick and apparently structureless with light microscopy, but electron microscopy shows that it consists of randomly oriented collagen fibrils. It is relatively resistant to invasion by pathogens and forms the anterior boundary of the stroma of the cornea,19,27 a laminated cross-ply structure which makes up 90% of the thickness of the cornea (0.7 mm peripherally and 0.53 mm centrally). It consists of more than 200 lamellae (layers or strips) of varying width and composed of collagen fibrils stacked one upon another and all running parallel to the corneal surface, each lamella being 1–2 µm thick. All fibrils within any given lamella are strictly parallel and are oriented at right angles to those in the lamella in front and in the lamella behind. Moreover, the fibrils are of uniform diameter (about 35 nm) and each fibril is separated from all its immediate neighbours by a more or less constant interval (about 50 nm); the space between the fibrils is filled by a ‘ground substance’ which consists of proteoglycans, glycoproteins and salts.
A number of explanations for the cornea’s transparency have been proposed over the past century and for a time it was attributed to the regular lattice-like arrangement of the collagen fibrils in human cornea. But the lack of this arrangement in other transparent tissues such as the dogfish cornea or the human lens points to another explanation. The cornea exhibits birefringence (the property of having more than one refractive index according to the direction of the transmitted light) and this supports the view that stromal collagen fibrils have a different refractive index from that of the stromal ground substance. The current understanding of corneal transparency28 is based on diffraction theory. This shows that scattering of light passing through a medium does not occur when variations in its refractive index extend over distances less than half the wavelength of the light. This is consistent with the diameter of the collagen fibrils found in the cornea and the spacing between them, both of which together are well below half the shortest wavelength of light (380 nm). In contrast with the corneal collagen, scleral collagen fibrils vary considerably in diameter (25–480 nm) and are grouped in tightly packed interweaving bundles of varying dimensions; also, the intervening spaces between bundles are large relative to the wavelength of light, all of which render the sclera opaque.
The uniform and close spacing of the collagen fibrils and their small diameter are thus fundamental to corneal transparency. The spacing is maintained by tight regulation of the hydration of the ground substance (mainly its glycosaminoglycans), collagen making up 15% of the cornea’s weight and water 78%. Dehydration or excessive hydration of the ground substance disturb the orderly arrangement of the fibrils and so impair the cornea’s transparency. As discussed above, the highly ordered laminar lattice-like configuration of the human cornea is not essential for its transparency. Its purpose, rather, is to maintain the cornea’s shape, curvature, and mechanical strength. It enables the cornea to withstand heavy impacts, though not always without damage to internal structures.
Because the degree of hydration of the stromal ground substance critically affects the spacing of the collagen fibrils and hence the optical properties of the corneal stroma, we should consider how this is regulated. In its normal transparent state the cornea is thin and relatively dehydrated, i.e. by about 20%, and its stroma is therefore hyperosmotic.19,29 The reason for this rests with the epithelium and the endothelium, particularly the latter. Both are semipermeable membranes acting as barriers to the diffusion of electrolytes and the flow of water across them. For the epithelium, the flow of water is from the tears, while for the endothelium from the aqueous fluid. The epithelium, having tight junctions (zonulae occludens) between its superficial cells, is more effective in this respect than the endothelium and prevents excessive absorption of water from the tears. The endothelium is a single layer of homogenous cells, about 5 mm thick, lining the internal surface of the cornea; damage to this layer is manifested by clouding of the cornea with an excess of fluid in the stroma. While being less of a barrier to the flow of water than the epithelium, the endothelium controls it by pumping excess water back into the aqueous fluid from where it came. Between the endothelium and stroma is another transparent membrane (Descemet’s) which is 10 µm thick. It is regarded as the basement membrane of the endothelium.
The endothelial metabolic pump is a chemical engine which actively transports electrolytes and with them water from the stroma into the aqueous. The principal ion involved is bicarbonate.30 This mechanism is temperature dependent and requires oxygen, glucose, carbohydrate metabolism, and adenosine triphosphatase; when deprived of any of these, corneal swelling occurs. Further evidence of the presence of this chemical engine comes from studies with metabolic inhibitors (poisons) such as ouabain.
Space does not permit a discussion of the structure, function and transparency of the natural lens. But it should be clear from this brief survey that both the cornea and the lens are marvels of precision in optical and biochemical engineering. Both are highly organized structures whose development and maintenance require a huge amount of information to be encoded in the genome. In particular, the function of the cornea as a lens in the case of man, is indispensable together with the neural pathway (retina to brain), for any vision more than bare perception of light.
The cornea is nourished by nutrients in the aqueous fluid via the endothelium and in the blood via capillaries at the limbus. It receives oxygen principally via the epithelium; oxygen is dissolved in the precorneal tear film, directly from the ambient air when the eye is open, and from blood passing through capillaries of the conjunctiva lining the inner surface of each eyelid when the eye is closed. The tears are thus essential as a vehicle of oxygen for the cornea. If the cornea is allowed to dry its health and transparency will invariably quickly suffer, leading to vascularisation and even perforation. This is why loss of, or absence of an upper eyelid, e.g. at birth, is an ophthalmic emergency and a major threat to the cornea. The epithelium and the anterior two thirds of the stroma are richly innervated with pain receptors. The epithelium has adrenergic receptors which are believed to regulate the metabolic activity of the epithelium. Thus the cornea is one of the most sensitive parts of the body, 400–600 times more sensitive than the skin. It is exquisitely sensitive to touch and to drying of the tear film. In clinical experience, a denervated cornea is very vulnerable even without trauma; the epithelium is prone to break down and healing is delayed.
The eyelids

Click here for larger view
Eyelids31 are beautifully designed devices (Figure 3) which move, like the cover of a roll-top desk, and conform to the shape of the eyeball to close it. The upper lid is the more important, being larger than the lower and is essential for the health of the cornea. The lids serve two important functions: protection of the eyeball and maintenance of the precorneal tear film, besides signaling the state of wakefulness and attention of a subject! The movement of the upper eyelids spreads the tear film evenly over the surface of the cornea which thereby receives oxygen and salts.
Each eyelid has four layers in two groups of two: the skin and orbicularis muscle for closing the lid form the anterior lamella. Behind them is the tarsal plate with its lining of conjunctiva which make up the posterior lamella.32 The skin of the eyelids is much thinner than that elsewhere on the face and concertinas easily to allow rapid unimpeded movements of the lids. The tarsal plate is a small sheet of fibrous tissue which reinforces and stiffens the lid for added protection and to give attachment for the muscles moving the lid. The margin of each eyelid has lashes to trap and repel foreign bodies, insects, etc. The tarsal glands (Meibomian), of which there are 20–30 in the lower lid and 30–40 in the upper, are embedded in each tarsal plate; they secrete an oil, contributing an important layer of the tear film.
The position and movement of the eyelids are controlled by a delicate balance between opposing muscles, namely the levator muscle which lifts the upper lid and the orbicularis which closes the lids. The levator is a thin flat muscle which extends back to the apex of the orbit and anteriorly it has two insertions:33 a) to the upper border of the tarsal plate via a sheet of smooth muscle (Müller’s) and b), via its aponeurosis (Greek apo, from + neuron, sinew, i.e. a fibrous sheet acting as a tendon for the muscle), at the same level to the connective tissue of the orbicularis.34 Its action is synchronised with the muscles moving the eyeball so that when the eye looks up the upper lid moves up also and vice versa when looking down; but for blinking it relaxes independently of the muscles moving the eye. The orbicularis is a thin sheet of muscle fibres which sweeps across each lid under the skin and parallel with the lid margins. Its fibres are thus concentrically arranged as arcs above and below the lid opening (the palpebral fissure); they are attached to bone (the orbital margin) and the lid ligaments at each end so that when they contract they act like a sphincter to close the lids. Normal gentle blinking involves the pretarsal muscle fibres, but with forceful blinking or spasm the fibres further away from the lid margins come into play.35
And so the eyelids clearly show abundant evidence of design in their structure. But, as with the visual pathway (retina to brain), their function is totally dependent at the same time on an intact coordinated nervous control system centred in the brain. An eye that will not open is blind and one that cannot close will rapidly fail from drying of the cornea. All has to be in working order for the eye to serve its purpose—another example of Behe’s irreducible complexity, requiring a considerable input of genetic information.
Reflex (involuntary) blinking with a latent period of only about 50 milliseconds occurs in response to many kinds of stimulation as a protective mechanism for the eye. The stimuli range from unexpected flashes of bright light, to sudden movement in the periphery of the visual field, to irritants of the eye and face, or to abrupt loud sounds.

Click here for larger view
Tears are secreted mainly by the lacrimal gland which is under nervous control and is located in a smooth wide depression in the upper outer corner of the orbit anteriorly. From there they are swept and spread by eyelid movements over the cornea towards the inner angle of the eye from where they drain into the nasal cavity (Figure 4). The drainage apparatus consists of two canaliculi (each a fine tube at the inner end of each lid) which converge to open into the lacrimal sac; the sac drains below into the nasolacrimal duct which opens in the nasal cavity. Part of the orbicularis muscle is involved in tear drainage into the nasal cavity by a pumping action36,37 which is lost when the muscle is paralysed with consequent overflow of tears onto the cheek. This pump is effective even when the head is inverted and has two phases, active and passive. The active phase occurs when the eye is closed by contraction of the orbicularis. At the same time groups of orbicularis muscle fibres which lie alongside each canaliculus contract and compress both the canaliculus and the lacrimal sac. In this way tears are driven down the nasolacrimal duct which has mucosal valves to prevent reverse flow, e.g. when blowing one’s nose. During the passive phase tears are drawn into each canaliculus by capillary attraction and by negative pressure generated by elasic recoil of the canaliculi and the sac.38,39
Tears
The tears40,41 form a film that covers and adheres to the cornea and the conjunctiva (the surface layer over the sclera anteriorly and lining the inner surface of the eyelids). This film, principally as a carrier of oxygen, is vital for the health of the cornea and for the refraction of light entering the eye. The tears also lubricate the constant frictional movements between the eyelids and the eyeball, remove exfoliated epithelial cells and irritants, and carry antimicrobial proteins.
As mentioned above, tears are secreted mostly by the lacrimal gland,42 i.e. their aqueous element and the importance of its nervous control has been underlined from clinical experience, in that operations to denervate the gland as a treatment for a watering eye resulted in a profound reduction of tear production.43 Such operations were abandoned because patients were left with a dry eye, a far worse condition. 8–12 ductules convey tears from the main gland and open in the upper conjunctival fornix (Latin, arch or vault) laterally.
The tear film, about 7 µm thick, is a remarkably stable structure which remains intact for at least 30 seconds in young normal subjects before breaking up while staring. It thus fulfils the definition of a film, namely a thin fluid layer that resists gravitational flow when vertical. An important factor for the stability and adhesion of the film to the cornea and conjunctiva is the microfolding (microplicae) of the surface of the corneal and, to a lesser extent, the conjunctival epithelia. In histological section, the microplicae appear as microvilli (microscopic finger-like projections).40 When this surface folding is damaged, e.g. with chemical burns or viral infections, the film loses its stability and tends to break up easily. The film comprises three major components in two layers: an oily secretion, aqueous secretion and mucin. The oil forms a very thin outer layer (about 0.1 µm deep) and is secreted by the Meibomian glands embedded in the tarsal plate of each eyelid. Expulsion of oil from these glands is aided by the muscular action of blinking. This oily layer of the tear film retards evaporation of the aqueous element of tears. Normally, when tear production is not raised by emotional44 or reflex stimulation, it acts a barrier to prevent the overflow of tears onto the cheek. The oil would not spread over the aqueous secretion without surfactants, mainly proteins. This contributes to the stability of the tear film.45
The inner layer (about 7 µm deep) forms the bulk of the tear film and current opinion46 suggests it is a mucin-dominated gel, hydrated by aqueous fluid containing electrolytes, small organic compounds and many proteins. The mucin element (glycoproteins) is secreted by scattered specialised epithelial cells of the conjunctiva and possibly also the corneal epithelium. Human tears have a low surface tension compared with water, because of their surface-active agents, but have a high viscosity. Their mucin is thought to be responsible for their non-Newtonian viscosity characteristics, i.e. their viscosity falls in conditions of high shear; this would explain in part the stability of the tear film during steady fixation between blinks but its minimal drag with lid or eyeball movements.45
The aqueous fluid, secreted by the lacrimal gland, contains a host of antimicrobial proteins46 which are the focus of continuing research. Similar proteins are also present in other mucosal secretions which are all products of the common mucosal defence system,47 comprising the alimentary, respiratory and genitourinary tracts, the mammary glands and the eye. Surface immunization to pathogens is induced in subepithelial lymphoid tissue (e.g. that of Peyer’s patches of the small intestine, the appendix, the tonsils and adenoids) which samples material within the lumen of the gut and respiratory tracts. This triggers the cloning of specialised lymphocytes producing specific antibodies, the lymphocytes travelling via lymphatics and the blood stream to seed e.g. the lacrimal and mammary glands. The variety of antibody produced by these particular lymphocytes is called secretory immunoglobulin A (IgA). Antibodies of this type are well suited to the mucosal environment because, unlike other classes of antibodies, they are resistant to various proteolytic enzymes. Besides secretory IgAs, the lacrimal glands secrete nonspecific antibacterial proteins such as lysozyme (a bacteriolytic enzyme), lactoferrin (an iron-binding protein impeding bacterial metabolism), cystatin (a bacterial proteinase inhibitor) and recently discovered in tears, lipocalin, another proteinase inhibitor. If injury or infection occurs, inflammatory cells from the blood are conveyed via the tear fluid.
Summary and conclusions
This brief review has perhaps given a glimpse of some of the human visual system’s superlative optical, mechanical and coordinated biochemical engineering. We tend to take it for granted but its construction requires enormous amounts of information to be stored in the genome. This must raise the question, Where does this information come from? Dawkins believes that, given sufficient time and generations, this information could have accumulated spontaneously in small increments through mutations and natural selection. However, it is certain not to have come through mutations since there is no observational or experimental evidence that they have ever added genetic information, rather the opposite.
We have also considered, in relation to the human eye, a number of examples of irreducible complexity: the highly organized structure and the finely balanced biochemical mechanisms of the cornea; the dependence of the cornea on an adequate supply of the specialized fluid, the tears, and on the presence of the eyelids. In addition, both the lids and the secretion of tears require their respective nervous control systems if the eye is to fulfil its purpose. The eye continues to be a thorn in the flesh of evolutionists and to reveal the emptiness of their sophistry.
References
- Dawkins, R., The Blind Watchmaker, Penguin Books, London, 1991. Return to Text.
- Wieland, C., Seeing back to front: are evolutionists right when they say our eyes are wired the wrong way? Creation 18(2):38–40, 1996. Return to Text.
- Anon., An eye for creation: an interview with eye disease researcher Dr G. Marshall, University of Glasgow, Scotland, Creation 18(4):19–21, 1996. Return to Text.
- Gurney, P., Our inverted retina—is it really bad design ? Journal of Creation 13(1):37–44, 1999. Return to Text.
- Hitching, F., The Neck of the GiraffeorWhere Darwin Went Wrong, Pan, London, 1982. Return to Text.
- Spetner L.M., Not by Chance, The Judaica Press, Inc., New York p. 60, 1998. Return to Text.
- Behe, M.J., Darwin’s Black Box, Simon & Schuster, New York p. 39, 1998. Return to Text.
- Burgess, S., Hallmarks of Design, Day One Publications, Epsom, p. 10, 2000. Return to Text.
- Sarfati, J., Origin of life: the chirality problem, Journal of Creation 12(3):263–266, 1998. Return to Text.
- Sarfati, J., Origin of life: instability of building blocks, Journal of Creation 13(2):124–127, 1999. Return to Text.
- Sarfati, J., Hydrothermal origin of life? Journal of Creation 13(2):5–8, 1999. Return to Text.
- Spetner, Ref. 6, p. 160. Return to Text.
- Ross R.D., Is perception of light useful to the blind patient? Arch. Ophthalmol. 116: 236–238, 1998. Return to Text.
- When cataract (opacity of the lens) is so gross that all view of the retina is obscured, testing for light projection is an important preliminary before embarking on surgery. Defective light projection, in the absence of optic nerve or cerebral disorder, generally indicates severe retinal dysfunction and a poor prognosis for surgery. Return to Text.
- Shortsightedness (myopia) means that parallel incident light rays come a focus in front of the retina, i.e. the eyeball is too long, resulting in poor distance vision. Return to Text.
- Removal of an opaque lens (cataract) is normally followed nowadays in the same operation by implantation of an artificial one. Previously, simple removal of the lens was sufficient to restore adequate vision for many in the third world with the very basic lifestyle of a peasant and unable to afford spectacles. Return to Text.
- Katz, M., The human eye as an optical system; in: Tasman, W. and Jaeger, E.A. (Eds), Clinical Ophthalmology, Lippincott-Raven, New York, vol. 1, ch. 33. 1998. Return to Text.
- Regarding the human eye, let alone any other topic covered in The Blind Watchmaker, Dawkins makes several erroneous statements which have been left uncorrected in later editions. He says (p. 93) of the human optic nerve that it has three million nerve fibres whereas it actually has about one million; the same error appears on p. 17 (three million ganglion cells). The ganglion cells are said by Dawkins (p. 17) to ‘constitute the “electronic interface” between the photocells and the brain’ and to ‘preprocess the information … before relaying it to the brain’. In fact, however, processing of signals from the photoreceptors is much more (if not entirely) a function of cells in the retinal layers between the photoreceptors and the ganglion cells rather than the ganglion cells themselves. His diagram of the eye (p. 16) also has a number of errors. Yet another (p. 301) is where he says the retina has 125 million ‘colour-coding photocells’, whereas there are about 6.5 million of such (cones) while the rest of the photoreceptors are the 125 million or so rods for colour-free night vision. Dawkins is unrestrained in his castigation or ridicule of others’ errors or what he regards as such (e.g. p. 79); he would do well to attend to his own. Return to Text.
- Watsky, M.A., Olsen, T.W. and Edelhauser, H.F., Cornea and Sclera; in: Tasman and Jaeger, Ref. 17, vol. 2, ch. 4. Return to Text.
- Mishima, S. and Hedbys, B.O., Physiology of the cornea, Int. Ophthalmol. Clin. 8:527. Cited by Simons, K.; in: Tasman and Jaeger, Ref. 17, vol. 4, ch. 16A. Return to Text.
- Dua, H.S., Gomes, J.A.P. and Singh, A., Corneal epithelial wound healing, Br. J. Ophthalmol. 78:401–408, 1994. Return to Text.
- Marshall, G.E., Konstas, A.G.P. and Lee, W.R., Collagens in ocular tissues, Br. J. Ophthalmol. 77:515–524, 1993. Return to Text.
- The existence and importance of the limbal stem cells in their task of continually replacing corneal epithelial cells has only comparatively recently become apparent. Previously it had not been fully appreciated how specialised the corneal epithelium is and how essential it is for corneal transparency. Depletion of the stem cell population from injury, e.g. chemicals or radiation, results in overgrowth of the cornea by conjunctiva with consequent impairment of vision and failure of corneal grafts. Limbal stem cell autografts have transformed the prospects for eyes so injured. See also Sarfati, J., ‘Stem cells & Genesis’, this issue, pp. 19–26. Return to Text.
- Dua, H.S., Stem cells of the ocular surface: scientific principles and clinical applications, Br. J. Ophthalmol. 79:968–969, 1995. Return to Text.
- Dua, H.S. and Gomes, J.A.P., Clinical course of hurricane keratopathy, Br. J. Ophthalmol. 84:285–288, 2000. Return to Text.
- Dua, H.S., Gomes, J.A.P. and Singh, A., Corneal epithelial wound healing, Br. J. Ophthalmol. 78: 402, 1994. Return to Text.
- Smolek, M.K. and Klyce, S.D., Cornea; in: Tasman and Jaeger, Ref. 17, vol. 1, ch. 8. Return to Text.
- Freegard, T.J., The physical basis of transparency of the normal cornea, Eye 11: 465–471, 1997. Return to Text.
- Van Meter, W.S., Holland, E.J. and Doughman, D.J., Corneal oedema; in: Tasman and Jaeger, Ref. 17, vol. 4, ch. 16A. Return to Text.
- Hodson, S.A. and Sherrard, E.S., The specular microscope: its impact on laboratory and clinical studies of the cornea, Eye 2:Suppl S85, 1998. Return to Text.
- Records, R.E., The eyelids; in: Tasman and Jaeger, Ref. 17, vol. 2, ch. 1. Return to Text.
- Shore, J. W., Mccord, C. D., Popham, J. K. Surgery of Eyelids; in: Tasman and Jaeger, Ref. 17, vol. 5, ch. 78. Return to Text.
- Werb, A., Senile ptosis, Trans. Ophthalmol. Soc. UK, 104:22–25, 1985. Werb’s anatomical description of the levator aponeurosis differs from that found in standard textbooks; it is based on his cadaver and histological studies besides his observations in clinical and surgical practice. I have personally confirmed his conclusions clinically and during surgery. Return to Text.
- There are racial variations of upper lid anatomy; the diagram shown in this article represents what is typical of the Caucasian variant. In the upper lid of Mongolians, the levator aponeurosis is inserted at a lower level, near the lid margin, so that the lid has a fuller appearance with a lower skin crease. Jeong et al., The Asian upper eyelid: an anatomical study with comparison to the Caucasian eyelid, Arch. Ophthalmol. 117: 907–912, 1999. Return to Text.
- A recent paper has emphasized the importance of a bundle of the orbicularis fibres which runs along the lid margin and is principally responsible for lid closure with normal blinking. Mackie, I.A., Riolan’s muscle: action and indications for botulinum toxin injection, Eye 14:347–352, 2000. Return to Text.
- Tanenbaum, M. and McCord, C.D., The lacrimal drainage system; in: Tasman and Jaeger, Ref. 17, vol. 4, ch. 13. Return to Text.
- Doane, M.G., Blinking and the mechanics of the lacrimal drainage system, Ophthalmology 88:844–851, 1981. Return to Text.
- The tears form a meniscus along the margin of each eyelid and flow from the outer angle of the eye towards the inner angle where a small pool of tears (the lacus lacrimalis) gathers. The punctum (orifice) of each canaliculus dips into this ‘lake’. During lid closure the two puncta ‘kiss’ and prevent regurgitation of tears as the canaliculi are compressed. When the lids separate the puncta ‘pop’ apart and suck tears out of the lake. Return to Text.
- In passing we may note that, contrary to the assertions of evolutionists, the plica semilunaris (Latin: plica, a pleat or fold) is not a vestigial functionless curiosity, a relic of the nictitating membrane found in animals. Its purpose is to enable unrestricted mobility for the eyeball when abducted (turned outwards). The conjunctiva is the surface membrane lining the eyelids and covering the anterior part of the sclera; to allow the eyeball and lids to move independently it forms a continuous pouch or sac above, laterally and below. But medially, because of the presence of the lacrimal drainage apparatus, there is no conjunctival sac; instead there is the plica semilunaris which is a crescentic fold of conjunctiva. It arises in the upper fornix towards its medial end, extends downward, concentric with the limbus, to end in the medial third of the lower fornix. When the eye is abducted the plica partially unfolds as the conjunctiva stretches so that movement is unimpeded. When the eye is adducted (turned inwards) a fibrous extension from the sheath of the muscle contracting (the medial rectus) draws the plica posteriorly, partially unfolding it and deepening the lacus lacrimalis. See Records, R.E., The conjunctiva; in: Tasman and Jaeger, Ref. 17, vol. 2, ch. 2. Return to Text.
- Records, R.E., The tear film; in: Tasman and Jaeger, Ref. 17, vol. 2, ch. 3. Return to Text.
- Bron, A.J., The Doyne Lecture, Reflections on the Tears, Eye 11:583–602, 1997. Return to Text.
- A number of small accessory or supplementary lacrimal glands, also under nervous control, are also present in the upper and lower fornices of the conjunctiva. Return to Text.
- Wright, P., Normal tear production and drainage, Trans. Ophthalmol. Soc. U.K. 104:351, 1985. Return to Text.
- Weeping for emotional reasons only occurs in humans. Return to Text.
- Bron, Ref. 41, p. 586. Return to Text.
- Gachon, A-M.F. and Lacazette, E., Tear lipocalin and the eye’s front line of defence, Br. J. Ophthalmol. 82:453–455, 1998. Return to Text.
- McClellan, K.A., Mucosal Defense of the Outer Eye, Surv. Ophthalmol. 42:233–246, 1997. Return to Text.
Readers’ comments
Comments are automatically closed 14 days after publication.