

Journal of Creation 22(2):67–74, August 2008
Browse our latest digital issue Subscribe
Did eyes evolve by Darwinian mechanisms?
The evolution of the eye has always been a dilemma for evolutionists from Darwin’s time to the present. Although Darwin, Richard Dawkins and other evolutionists have tried to explain how an eye could evolve, their solutions are clearly unsatisfactory. Many kinds of eyes exist, but no progression of eye designs from simple to complex can be produced in the natural or fossil world. Furthermore, the simplest ‘eye’, the eyespot, is not an eye but pigmented cells used for phototaxis; yet even it requires an enormously complex mechanism in order to function as a vision system.
The concept of irreducible complexity (IC) has become an important tool in intelligent design theory. One of the best examples of IC is the design of the animal eye. Eyes are critical because, for the ‘vast majority of animals’, vision is their ‘most important link to the world’.1 Darwin vividly recognized the problem of eye evolution and the serious impediment that it was for his theory. In his words,
‘To suppose that the eye, with all its inimitable contrivance for adjusting the focus to different distances, for admitting different amounts of light, and for the correction of spherical and chromatic aberration, could have been formed by natural selection, seems, I freely confess, absurd in the highest possible degree.’2
Nonetheless, Darwin felt the seemingly insurmountable problem of the evolution of what he called an organ of ‘extreme perfection and complication’ could be solved.2 He included a three-page proposal of intermediate stages through which eyes might have evolved via gradual steps.3 These stages included the following:
- photosensitive cell
- aggregates of pigment cells without a nerve
- an optic nerve surrounded by pigment cells and covered by translucent skin
- pigment cells forming a small depression and then a deeper depression
- the skin over the depression gradually taking a lens shape
- evolution of muscles that allow the lens to adjust.
These stages in living animals are believed to constitute major evidence for the evolution of the eye.4 Isaak claims that all of these steps are viable because all of them exist in animals living today:
‘The increments between these steps are slight and may be broken down into even smaller increments. Natural selection should, under many circumstances, favor the increments. Since eyes do not fossilize well, we do not know that the development of the eye followed exactly that path, but we certainly cannot claim that no path exists.’5
University of Chicago biology Professor Jerry Coyne wrote that human
‘ … eyes did not suddenly appear as full-fledged camera eyes, but evolved from simpler eyes, having fewer components, in ancestral species. Darwin brilliantly addressed this argument by surveying existing species to see if one could find functional but less complex eyes that not only were useful, but also could be strung together into a hypothetical sequence showing how a camera eye might evolve. If this could be done—and it can—then the argument for irreducible complexity vanishes, for the eyes of existing species are obviously useful, and each step in the hypothetical sequence could thus evolve by natural selection.’6
The dominant theory was outlined by Dennett, who concluded that all eye evolution requires is a
‘ … rare accident giving one lucky animal a mutation that improves its vision over that of its siblings; if this improvement helps it to have more offspring than its rivals, this gives evolution an opportunity to raise the bar and ratchet up the design of the eye by one mindless step. And since these lucky improvements accumulate—this was Darwin’s insight—eyes can automatically get better and better and better, without any intelligent designer.’7
Others are not so confident. Melnick concluded that the eye is a marvel and that ‘its immense complexity and diversity in nature, as well as its beauty and perfection in so many different creatures of the world, defies explanation even by macroevolution’s most ardent supporters.’8 This paper explores these conflicting views.
Evolution of the eye
Advanced vision appears almost at the very beginning of the fossil record. The oldest eye in the fossil record, that of a trilobite, is a very complex faceted compound eye that ‘dates’ back to the Cambrian, conventionally dated about 540 million years ago.9,10 The fossil evidence shows that from the beginning of the fossil record eyes are very complex, highly developed structures. We also have ‘living fossils’, animals that have remained virtually unchanged since very early in history. University of Salford biologist, Laurence R. Croft, wrote that the ‘precise origin of the vertebrate eye is still a mystery. The fascinating thing about the evolution of the eye is its apparent sudden appearance.’11 Specifically, the fossils show that vision originated ‘in the early Cambrian’, which Darwinists put at ‘some 530 million years ago’.12
Furthermore, although the ‘Cambrian animals were not the same species as exist today … nearly all the modern phyla had rapidly come into existence, fully equipped with eyes as far as can be told from the fossils’ and during the Cambrian explosion ‘something remarkable seems to have happened … a rich fauna of macroscopic animals evolved, and many of them had large eyes.’12 Sir Steward Duke-Elder, the preeminent ophthalmologist at the time of his death in 1979, acknowledged the sudden appearance of the perfected vertebrate eye, noting:
‘The curious thing, however, about the evolution of the vertebrate eye is the apparent suddenness of its appearance and the elaboration of its structures in its earliest known stages. There is no long evolutionary story as we have seen among invertebrate eyes, whereby an intracellular organelle passes into a unicellular and then a multicellular eye, attaining by trial and error, along different routes an ever-increasing degree of complexity. Within the vertebrate phylum the eye shows no progress of increasing differentiation and perfection as is seen in the brain, the ear, the heart and most other organs. In its essentials the eye of a fish is as complex and fully developed as that of a bird or man [emphasis added].’13
Biochemical studies have shown that the human lens contains
‘ … proteins similar to those found in the cyclostomes (hagfishes and lampreys) that are the living descendants of the Agnatha, which originated the vertebrates about 450 million years ago. Thus these studies have confirmed the view that the vertebrate eye, and in particular the lens, has changed very little during the course of evolution.’14
Evidence for eye evolution from living animals
Only about a third of all animal phyla contain species with proper eyes, another third contain species with light-sensitive organs only, and a third have no means of light detection, although many can detect heat.15 Nonetheless, of those animals with eyes, both vertebrates and most invertebrates, an enormous variety of eye designs, placement and sizes exists.10 The eyeball diameter ranges from less then a tenth of a millimetre in certain water fleas to 370 mm in the giant squid.16 Eye placement also varies, ranging from the common binocular vision employed by most mammals to the movable eye on each side of the head used by many lizards.
The number of eyes in one animal can also vary from none to eight. In spiders alone the number ranges from zero to eight, always existing in pairs of two. Some eyes contain both a lens and a retina-like structure in a single cell.17 A complex telephoto lens was identified in the chameleon in 1995. The reason why so many designs exist is because eyes must serve very different life forms that live in very different environments. Animals live in the ground, inside of other animals, in the air, on land, in salt water and in fresh water. Furthermore, animals range in size from a water flea to a whale.
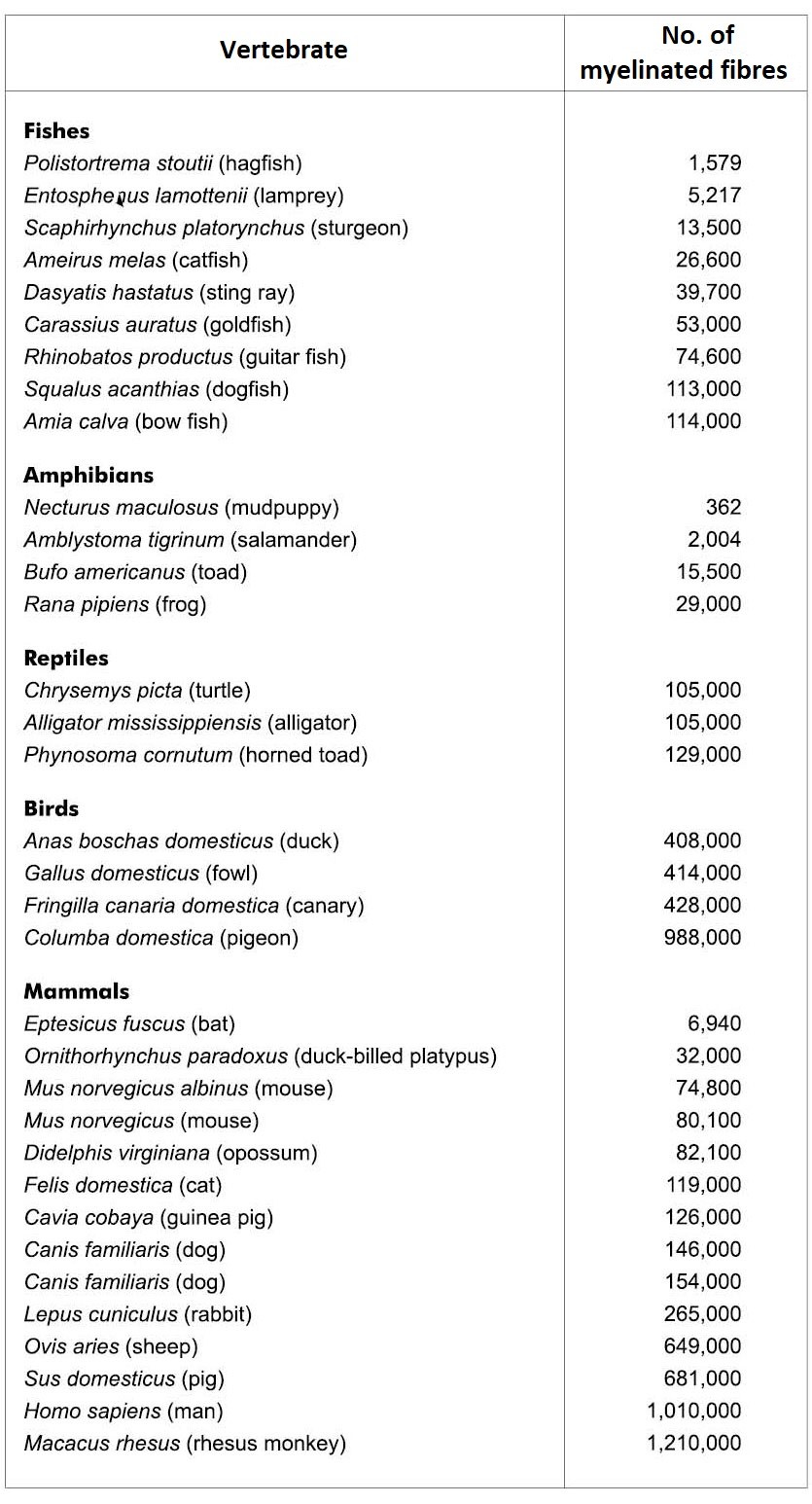
Although many kinds of very different eyes are known, no direct evidence exists to support the evolution of the eye and its accessory structures. Furthermore, much evidence contradicts such evolutionary beliefs. For example, note in table 1 that the number of myelinated fibres in the optic nerve does not correlate with putative evolutionary development. A pigeon has almost as many fibres as a human. Many birds, such as the eagle and hawk, have excellent vision yet have half as many fibres as a domestic pig.
Another example is visual pigments. The presumably highest, most evolved form of life, the higher primates, have only two cone photoreceptors, blue and green, but birds have a total of six pigments: four cone pigments plus pinopsin (a pineal photoreceptive molecule) and rhodopsin for black and white vision.12,18 Put another way, chickens, humans and mice all have the rhodopsin pigment; mice in addition have blue and green; humans have blue, green, and red; and birds have these three pigments plus violet and pinopsin. For every colour that humans perceive, birds can see very distinct multiple colours, including ultraviolet light. Birds use infrared light (which we sense as heat) for night vision, allowing them to rapidly visualize their young in a dense, dark tree.
The possibility of classifying eyes in living animals from simple to complex—simple types existing in simple animals and complex types in complex animals (which we will show cannot be done)—does not provide evidence for an evolutionary relationship. A primary problem is that this attempt is based only on eye characteristics as they presently exist. Historical eye evolution cannot be proven by listing a series of existing eyes from simple to complex and then arguing that the complex evolved from the simple because evolution requires that all existing eyes have an equally long evolutionary history.
According to neo-Darwinism, the simplest modern eye in living animals has had the same amount of time and evolutionary history as the most complex eye because life began about 3.5 billion years ago and all life today evolved from this point in history. Although Darwinists argue that many of these eyes are evolutionary dead ends, this would require an admission that these modern ‘simple’ eyes are only analogues or ‘similar’ to putative past ancestral eyes (to more complex modern types), which reduces their value as evidence.
Darwinists need to determine the eye designs from which existing eyes have actually descended, one from the other, over time. Duke-Elder and Darwin (1872) before him were unable to do this, yet they offered their list of eyes of varying complexity as evidence of evolution. Cousins wrote:
‘ … the crucial importance of this requirement to the theory of evolution was fully understood by Darwin, who stated that, in searching for the gradations through which an organ in any species has been perfected, we ought to look at its lineal progenitors. Indeed we ought; though he himself could not do so. It is deceptive to the reader to create a seriation beginning with eye spots as seen in unicellular organisms and call them, as does Duke-Elder (1958), the earliest stage of evolution.’19
Croft concluded that the claim that we can line up eyes in an evolutionary sequence from very simple to very complex is false because research on the developmental history of the eye in widely differing species finds
‘ … it remarkably similar. Indeed the basic features of the eye in different vertebrates are very much the same despite great variations in their mode of life and adaptation to habitat. Furthermore, unlike other organs such as the heart, there is no long evolutionary history with the eye. In essence the eye of a newt is as complex and fully developed as that of a man.’11
Sinclair also concluded that vertebrates and most invertebrates, including insects and cephalopods (molluscs, including octopuses and squid), all have eyes with common visual elements, including ‘a similar photoreceptor design’, yet have a marked ‘dissimilarity of their appearance’.10
The source of the design and evolution of the eye, Darwinists postulate, was a series of beneficial mutations that had to occur in appropriate unison in order to produce the set of structures required for eyes to function. The new mutation set, Darwinists argue, resulted in a superior structure compared to the old one, and this new and better eye improved the animals’ ability to compete against other forms of life. Some of the many problems with this conclusion were noted by Grassé in his discussion of Myrmelion (ant lion) anatomy:
‘Have you ever seen a mutation simultaneously affecting two separate components of the body and producing structures that fit one another precisely? … have you ever beheld three, four or five simultaneous mutations with matching structures producing coordinating effects? … These are vital questions that demand an answer. There is no way of getting around them, or evading the issue. Every biologist who wants to know the truth must answer them, or be considered a sectarian and not a scientist. In science there is no “cause” to be defended, only truth to be discovered. How many chance occurrences would it take to build this extraordinary creature [Myrmelion formicarius]’?20
An organ that did not aid the animal’s survival would use scarce energy, nutrients and body space and, if the organ were not used, would be at high risk for problems such as infection. An eye modification would not be selected until it was not only functional but produced a system demonstratively better than the existing organ. Only then could natural selection operate to choose from existing variations to perfect the organ beyond mere functional effectiveness.

Advanced eye designs
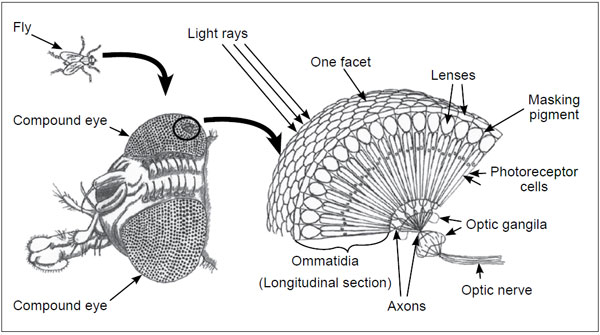
Many kinds of eyes exist, and there are many schemes to classify them. The most basic classification system groups all eyes into four classes. The first is the camera type or ‘simple’ eye, such as exists in humans, which uses a focusing system to project a single, sharp image on the retina. The second type is the fixed focus compound type (figure 1) that uses multiple separate refractive units called ommatidia, such as used by trilobites and flies. The third type is a scanning eye that builds an image much like a television camera, such as is used in the small marine crustacean copilia, which in females takes up more than half of its body.21 The fourth type is the complex eye, found in cephalopods and certain advanced vertebrata, consisting of a cornea, iris, lens, retina and numerous accessory structures.22
This division obscures many major differences: some shrimp have a combined simple and compound eye, which is actually a third basic eye type, not a transitional form. This division system also greatly oversimplifies the variety that exists because ‘at least eleven distinct optical methods of producing images’ are now known.23 The classification system used in this paper was developed by Land and Nilsson (2005) and is given in table 2 (see also figure 2).
Problems with classification

Note that the most logical classification of eye types is into some type of evolutionary classification from simple to more complex, but this list does not lend itself very well to a hierarchy as postulated by Darwin. Actually, arranging just the 10 basic eye designs used in the Land and Nilsson system from simple to complex is impossible. For example, types A, B, C, D, E, F, H, and I appear similar in complexity, and types G and J appear more complex but are found in lower forms of life (in some winged insects and crustaceans). In Land’s classification the ‘simplest’ type (A) and the most complex type (J), are both found in crustaceans (crustaceans use designs in groups A, E, F, G and H, and molluscs those in group A and H). Nearly identical optical designs are found in very ‘distinctly unrelated animals’ such as fish and cephalopods.24 The Land list groups the basic eye designs and optical systems only, ignoring the design of the retina cells, the many supportive cells, (such as the ganglion cells, amacrine cells, horizontal cells and bipolar cells), the other nervous system components, including the optic nerve, and the optical system-processing centre, such as the occipital lobe of the brain.
Using these criteria would create even more problems in attempting to produce a hierarchy because the processing system is always much more complex than the light collection system, placing all known eye systems at the upper level of Darwin’s scheme. Of course, Darwin was not aware of the vision system’s enormous complexity or variety, nor was he aware of the complexity of the many accessory systems and processing structures such as the brain.
The problems of producing a simple to complex hierarchy are illustrated by the fact that the ten types are also commonly arranged into four basic eye designs: the holochroal eye, the superposition eye, the schizochroal eye, and the human apposition compound eye. All of these basic eye designs require a system of focusing resolution, and a complex neurological processing system to enable the viewer to make sense of the large mass of constantly changing signals sent by the retina or other light sensitive cells via the optical nerve to the brain efficiently and rapidly.
‘Despite decades of research, we still have only limited understanding of how vision actually works’, making it difficult to produce both consistent classification schemes and hierarchies in an attempt to postulate a reasonable evolutionary phylogeny.23 We do have a fairly good understanding of the eye structure itself, which allowed construction of the classification above. Contrary to evolutionary expectations, the eyes of phylogenetically distant life forms can be very ‘similar in a large number of details.’16
Ironically, the greatest variety of eye design, not only in structure, but also in number and location, exists not among the vertebrates as Darwinism would expect, but among the so-called ‘primitive’ invertebrates.16 Invertebrates also have eyes that are, in some respects, superior to those of vertebrates. One example is the hemispherical eyes of most flies and other insects, which produce, unlike human and most vertebrate eyes, an image largely free of spherical distortion.25 Human eyes have significant peripheral image distortion, but spherical eyes form a sharp image in all directions. However, humans do not have sharp peripheral vision because this is the function of the central retina called the macula. Our peripheral vision is for the detection of light and movement which trigger the fixation reflex to turn the eyes toward the stimulus.
Another problem in the theory that eye designs represent an evolutionary sequence is that eyes from the three major phyla (vertebrates, arthropods and mollusca) arise from different tissues and are radically different.26 For this reason, evolutionists concluded that they have separate evolutionary histories, and the many similarities that exist are due to presumed evolutionary convergence.26 In essence, ‘we don’t know how it could possibly have evolved, so it must have evolved over and over.’ The eye differences would be due to the different needs and circumstances of each organism and its habitat, irrespective of any evolutionary connection. Yet another problem is the evidence for eye evolution forces the conclusion that most of these eye designs must have evolved ‘in a brief period during the Cambrian.’17
The simplest eye
Darwinists often claim the primate eye is the most evolved, but many mislabelled ‘primitive’ eyes have advantages over ours. For example, the human eye can register up to 60 images per second; a lowly bee about 300 per second. For this reason, bees can see far better while rapidly moving. The motion picture standard (24 frames per second), to a bee, would be viewed as a series of still pictures. For humans the frames are blurred, giving the illusion of motion. This design innovation in so-called primitive animals is more complex than the corresponding structure in the human eye.
The simplest eye type known is the ocellus, a multicellular eye comprising of photoreceptor cells, pigment cells and nerve cells to process the information—is step 4 in Darwin’s list.27 The most primitive eye that meets the definition of an eye is the tiny—about the size of the head of a pin—microscopic marine crustacean copepod copilia. Only the females possess what Wolken and Florida call ‘remarkable eyes which make up more than half of its transparent body.’28 Claimed to be a link between an eyespot and a more complex eye, it has two exterior lenses that raster like a scanning electron microscope to gather light that is processed and then sent to its brain.29 It has retinal cells and an eye ‘analogous to a superposition-type ommatidium of compound eyes’.30 This, the most primitive true eye known, is at stage 6 of Darwin’s evolutionary hierarchy!
Visual cell differences
Evolution would predict that the more advanced an eye, the more detail it can pick up, a factor related to the number of visual cells. This is not what is often found. In a ‘simple’ visual system (brain and retinas) the smallest number of visual cells is found in the plethodontid salamander, T. narisovalis, which uses about 65,000 cells for the entire visual brain centre and 60,000 for the retina alone. This ‘extraordinarily low’ number of cells is used not because the animal is primitive but because it has a very small head, eye, and brain plus relatively large cells.31 They add that the smallest extant salamander, T. pennatulus (which is much smaller than T. narisovalis), has about 94,000 visual cells and about the same number of retinal cells. For comparison, the brain visual centres of the frog S. limbatus contain about 400,000 cells. This illustrates the fact that evolution cannot be argued
‘ … by asserting that the eye can be built up gradually from a single patch of light-sensitive skin through various stages, slowly reaching the complexity of the vertebrate camera eye. …
the case for the evolution of the vertebrate eye or even a light-sensitive patch of skin … must be made in regard to the entire complexity of the living organism, at least insofar as that complexity supports vision (even in the least complex form). For this reason, the debate shouldn’t be about the evolution of the eye, but about the evolution of vision, and vision is always the vision of some particular kind of living animal, a living whole in which the integrated activity and experience of seeing, even in its simplest form, can take place.’32
In addition to number of cell differences, photoreceptor cell differences also exist. The cells that provide the membrane surface for opsin molecules can be either ciliary or microvillar structures. The microvilli type dominates in invertebrates, and ciliary types in vertebrates. Even physiological responses vary widely. Light causes microvillous receptors of arthropods and molluscs to depolarize but causes the ciliary receptors of vertebrates to hyperpolarize. Invertebrates use inositol triphosphate for photo-transduction in the second messenger system, whereas vertebrate photoreceptors use cyclic Guanosine 5’-Monophosphate (GMP). Although opsin is the key molecule used to detect light in both vertebrates and invertebrates, regeneration mechanisms (reisomerization) of the chromophore/opsin system ‘are dramatically different among phyla’.33 Other important differences include invertebrate eyes that are formed from the dermal surface of the ectoderm and vertebrate eyes that are formed from the neural ectoderm.34
Another problem for evolution is that at least 11 distinct optical methods are used to produce images. For one type to evolve into a more ‘advanced’ type ‘requires intermediate stages that are much worse or useless compared with the existing design. This would make a switch essentially lethal to animals that depend on sight.’35 For example, the advanced rods and cones in ‘primitive’ animals and the lack of evidence for their evolution has motivated some to conclude that the ‘basic tetrachomatic system evolved very early in vertebrate evolution.’36 Furthermore, no progression from simple to complex photoreceptors exists, but rather only ‘four spectrally distinct classes of cone pigment encoded by distinct opsin genes’ is found in the natural world.37
Evaluation of genes involved in eye development
Conversely, similarities, such as the fact that some of the genes involved in eye development are very similar in most animals, argue for a single evolution of the eye. Yet, the difficulties of eye evolution are so great that eyes are hypothesized by some researchers to have independently evolved at least 40 and as many as 65 times.38 As Fernald notes, at present, ‘we do not know whether eyes arose once or many times, and, in fact, many features of eye evolution are still puzzling.’23 A better explanation for the same gene being used by different animals (or plants) is for economy of design by a higher Intelligence.
Vertebrate eyes could not have evolved in isolation because eye parts do not have a function as self-contained entities. Eyes are part of very complex, interconnected living organisms, and eyes are only one part of the vision system.39
One gauge to help determine eye complexity is the number of genes involved in producing the eye—the more genes that are required, the more complex the eye may be. In the primitive Drosophila, so far 501 eye-related genes have been identified, or about 3.5% of its entire genome.24 Vertebrate eyes are estimated to involve 7,500 genes just to develop and regulate the retina—or about 30% of the entire human genome of 25,000 genes.24
Views on eye evolution have flip-flopped
These problems are part of the reason why ‘views on eye evolution have flip-flopped, alternately favoring one or many origins.’40 The markedly distinct ontogenetic origin of eyes in very different species is one reason why eyes are postulated to have evolved 40 or more times independently.40 For example, the eyes in many molluscs, including some cephalopods such as squids and octopuses, are remarkably similar to vertebrate eyes. Both have a cornea, a lens, an iris and a retina. One of the major differences is, in one, the retina is inverted, compared to the other.41
Evolutionists attempt to solve this problem by assuming that the phylogenetic line that led to molluscs split very early in evolutionary history, long before the eye had evolved. Then they postulate parallel evolution—concluding that the two eyes evolved to be almost identical, yet were completely independent of each other. Of note is the fact that the most ‘primitive’ camera eye known (the nautilus pinhole eye) and the most advanced eye known are both found in cephalopods! Molluscs as a group contain a pigment eyespot design, a pigment cup (cupulate), a simple optic cup with a pinhole lens, an eye with a primitive lens (a murex marine snail) and a complex eye (the octopus), the latter which is the ‘most elaborate’ eye in the invertebrate kingdom.42
Embryonic origin of vertebrate eyes in contrast to cephalopod eyes
Another major difference is found in the embryonic origin of many structures in vertebrate eyes in contrast to cephalopod eyes. For example, cephalopod eyes form from an epidermal placode by successive infoldings, whereas vertebrate eyes develop from the neural plate, and the overlying epidermis forms the lens. Yet another problem for eye evolution is that the eye of just one evolutionary related class, the vertebrates, ‘develops from a diverse collection of embryonic sources through a complex set of inductive events.’43
Conclusions
Dennett wrote that the eye lens is ‘exquisitely well-designed to do its job, and the engineering rationale for the details is unmistakable, but no designer ever articulated it.’44 He concludes that its design is not real, but an illusion because evolution explains the eye without the need for a designer. This review has shown that evolution does not explain the existence of the vision system, but an intelligent designer does. The leading eye evolution researchers admit they only ‘have some understanding of how eyes might have evolved’.45 These explanations do not even scratch the surface of how a vision system could have arisen by evolution—let alone ‘when’.
Much disagreement exists about the hypothetical evolution of eyes, and experts recognize that many critical problems exist. Among these problems are an explanation of the evolution of each part of the vision system, including the lens, the eyeball, the retina, the entire optical system, the occipital lobes of the brain, and the many accessory structures. Turner stressed that ‘the real miracle [of vision] lies not so much in the optical eye, but in the computational process that produces vision.’46 All of these different systems must function together as an integrated unit for vision to be achieved. As Arendt concludes, the evolution of the eye has been debated ever since Darwin and is still being debated among Darwinists.47 For non-evolutionists there is no debate.
Acknowledgments
I would like to thank Curt Deckert, Clifford Lillo, Eric Belivert, Terra Richmond DO, John UpChurch and Jody
Allen, for their help.
Re-posted on homepage: 1 December 2017
References and notes
- Sinclair, S., How Animals See: Other Visions of Our World, Henry Holt, New York, p. xi, 1985. Return to text.
- Darwin, C., The Origin of Species, John Murray, London, p. 186, 1859. Return to text.
- Darwin, C., The Descent of Man, and Selection in Relation to Sex, John Murray, London, 1872. Return to text.
- see Shermer, M., The Case Against Intelligent Design, Henry Holt, New York, p. 17, 2006. Return to text.
- Isaak, M., The Counter-Creationism Handbook, Greenwood Press, Westport, CT, p. 65, 2005. Return to text.
- Coyne, J., The faith that dare not speak its name: the case against intelligent design, The New Republic, p. 30, 2005. Return to text.
- Dennett, D.C., The hoax of intelligent design and how it was perpetrated in Brockman, (Ed.), Intelligent Thought: Science Versus the Intelligent Design Movement, Vintage Books, New York, p. 37, 2006. Return to text.
- Melnick, J., Vision: an evolutionary enigma, Christian Citizen 1(9):26, 1981. Return to text.
- Moses, K., Fly eyes get the whole picture, Nature 443:638–639, 2006. Return to text.
- Sinclair, S., ref. 1, p. 9. Return to text.
- Croft, L.R., The Last Dinosaurs, Elmwood Books, Chorley, Lancashire, p. 57, 1982. Return to text.
- Land, M.F. and Nilsson, D.-E., Animal Eyes, Oxford University Press, New York, p. 1, 2005. Return to text.
- Duke-Elder, S.S., System of Ophthalmology. Volume 1: The Eye in Evolution, The C.V. Mosby Company, St. Louis, p. 237–238, 1958. Return to text.
- Croft, ref. 11, p. 59. Return to text.
- Land and Nilsson, ref. 12, p. 4. Return to text.
- Land and Nilsson, ref. 12, p. 15. Return to text.
- Jager, S., Lewis, J., Zvyaga, T., Szundi, I and Sakmar, T., Chromosome structural changes in rhodopsin from nanoseconds following pigment photolysis, Proceedings of the National Academy of Science 94:8557–8562, 1997. Return to text.
- Cousins, F.W., The Anatomy of Evolution, Duffett Publications, London, p. 125, 2003. Return to text.
- Grassé, P.P., Evolution of Living Organisms, Academic Press, New York, NY, p. 163, 1977. Return to text.
- Wolken, J. and Florida, R.G., The eye structure and optical system of the crustacean capecod, copilia, The Journal of Cell Biology, 40(1):279–285, 1969. Return to text.
- Arendt, D. and Wittbrodt, J., Reconstructing the eyes of urbilateria, Philosophy Transactions of the Royal Society of London 356:1546, 2001. Return to text.
- Fernald, R.D., The evolution of eyes, Karger Gazette 64:2–4,
2001; p. 2. Return to text. - Fernald, R.D., Casting a genetic light on the evolution of eyes, Science 313:1914–1918, 2006; p. 1914. Return to text.
- Sinclair, ref. 1, p. xvii. Return to text.
- Fernald, R.D., The evolution of eyes, Brain, Behavior and Evolution 50(4):253, 1997. Return to text.
- Arendt and Wittbrodt, ref. 22, p. 1546. Return to text.
- Wolken and Florida, ref. 21, p. 279. Return to text.
- Sinclair, ref. 1, p. xiii. Return to text.
- Wolken and Florida, ref. 21, p. 280. Return to text.
- Roth, G. and Blanke, J., Vertebrates in McGraw-Hill Yearbook of Science and Technology, McGraw-Hill, New York, 1997. Return to text.
- Wiker, B. and Witt, J., A Meaningful World: How the Arts and Sciences Reveal the Genius of Nature, InterVarsity Press, Downers Grove, IL, p. 44, 2006. Return to text.
- Fernald, ref. 16, p. 146. Return to text.
- Cousins, ref. 19, p. 284. Return to text.
- Fernald, ref. 23, pp. 2–3. Return to text.
- Bowmaker, J.K. and Hunt, D.M., Evolution of vertebrate visual pigments, Current Biology 15(13):R484–R489, 2006; p. R489. Return to text.
- Bowmaker and Hunt, ref. 36, p. R484. Return to text.
- Fernald, ref. 16, p. 145. Return to text.
- Breidach, O. and Kutsch, W., The Nervous Systems of Invertebrates: An Evolutionary and Comparative Approach. With a coda written by T.H. Bullock, 1995. Return to text.
- Fernald, ref. 24, p. 1917. Return to text.
- McIlwain, J.T., An Introduction to the Biology of Vision, Cambridge, New York, 1996. Return to text.
- Lovicu, F. and Robinson, M.L., Development of the Ocular Lens, Cambridge University Press, New York, p. 17, 2004. Return to text.
- Fernald, ref. 23, p. 3. Return to text.
- Dennett, D.C., Breaking the Spell: Religion as a Natural Phenomenon, Viking, New York, p. 60, 2006. Return to text.
- Fernald, ref. 16, p. 467, emphasis mine. Return to text.
- Turner, J.S., The Tinker’s Accomplice: How Design Emerges from Life Itself, Harvard University Press, Cambridge, MA, p. 161, 2007. Return to text.
- Arendt, D., Evolution of eyes and photoreceptor cell types, International Journal of Developmental Biology 47:563, 2003. Return to text.
- Mitchell, L., Mutchmor, J. and W. Dolphin, W., Zoology, Benjamin Cummings, Menlo Park, CA, p. 279, 1988. Return to text.
- Mitchell et al., ref. 48, p. 278. Return to text.
- Cousins, ref. 19, p. 300. Return to text.




Readers’ comments
Comments are automatically closed 14 days after publication.