

Journal of Creation 14(2):82–90, August 2000
Browse our latest digital issue Subscribe
Ecology, biodiversity and Creation
‘Lord, you have made many things …. All these things depend on you to give them their food at the right time.’1
Ecology, the study of natural relationships, has revealed numerous essential ‘biodiversity services’ without which, ecosystems could not function in their present form. Biodiversity studies emphasise the need for preserving intact ecosystems, but also unintentionally speak about the origin of ecology and even of the origin of life. The necessity of functioning ecosystems today suggests they would also have been needed in the past. If this is so, then a reasonable explanation for ecology is that living things were designed with the capacity to function within ecosystems.
Ecology is the branch of biology that attempts to understand relationships between organisms, and between them and their non-living surroundings. Ernst Haeckel coined the word, ‘ecology,’ in the 1860s.2 As a zoologist, and ardent follower of Charles Darwin, he hoped to find supporting evidence for evolution in ecology. The environment is understood to provide the selective forces in natural selection. Thus, understanding ecology seemed to be an important step in understanding evolution.
As relationships and conditions vary in a community, different selection pressures are imposed on its members. Thus, a community is dynamic with species varying over both space and time. Nevertheless, the concept of natural selection does not answer the question about how ecological relationships originated, except to invoke coevolution to provide them. It is supposed that as species evolved, so did ecology.
Coevolution is defined as:
‘… joint evolution of two or more non-interbreeding species that have a close ecological relationship; through reciprocal selective pressures, the evolution of one species in the relationship is partially dependent on the evolution of the other [emphasis added].’3
The problem is, since coevolution requires already existing ecological relationships, it cannot account for the origin of ecology.
It is possible for two species in close ecological relationship to refine their relationship through mutual selection, but this does not explain how they came to be ecologically related in the first place. There must be some other explanation.
On the contrary, accumulating evidence from ecology and biodiversity studies suggests something quite different from gradual evolutionary accumulation of species and step by step development of what would eventually become essential ecological relationships. The current indispensable nature of many ‘ecological services’, and the relationships that provide them, suggests that, just as ecological services are necessary now, past ecosystems would also have needed them, but not necessarily in identical ways. Moreover, the essential nature of ecological relationships now does not appear to allow time for evolutionary development of ecology. Ecosystems would have failed many times over without the full range of ecological services (see Figure 1).
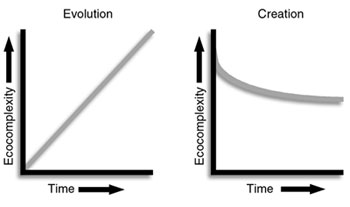
Behe, writing in Darwin’s Black Box,4 suggests that the complex nature of essential biochemical pathways in cells leaves little room for their gradual step-wise development, but rather requires that such pathways had to have begun as completely functioning systems. He thought this required design as an explanation for their origin. Javor writes: ‘It is reasonable, then, to suppose that when living cells were first brought into existence, all of their components must have been present and functioning. If this is so, then living cells had to be made rapidly.’ He then adds an important point: ‘The same suggestion may be made for all of the components of the ecological system, where mutual support and interdependence exist. It is sensible to suppose that these were created simultaneously.’5 Not only is there evidence for design of cells, but also, at the other end of the spectrum of life, for ecosystems.
It is a great irony that, as we examine complex and essential ecological relationships, ecology may turn out to be a significant challenge to the evolutionary ideas that ecological studies were first undertaken to support.
Biodiversity
The word, ‘biodiversity’, was first used in 1986 at a conference at the Smithsonian Institute in Washington, D.C.6 However, while the term may be new, the idea has been understood much longer, even going back to Darwin’s time.7
Since 1986, biodiversity has been the subject of a rapidly growing number of articles and books. Baskin refers to biodiversity as an emerging science.8 Two books I have found useful are: The Work of Nature: How the Diversity of Life Sustains Us, by Yvonne Baskin8 and Nature’s Services: Societal Dependence on Natural Ecosystems, edited by Gretchen C. Daily.9 I will cite from these works, and others. In general, The Work of Nature has been the most useful for this present paper.
Biodiversity refers to the collection of species in ecosystems, different populations of those species with their genetic variations (estimated to number as many as 220 populations per species for an estimated total of between 1.1 and 6.6 billion populations world-wide10), and especially the collective ecological services they provide. Taken together, these three entities produce an enormous amount of structural and functional variation and interdependence. Biodiversity, and the services it provides, may be compared to a jigsaw puzzle in which it is impossible to remove just one puzzle piece. Whenever you attempt to do so, you end up removing several other pieces. In like manner, ecological linkages between organisms in ecosystems make it difficult to remove just one species.
No organism lives independently, but both gives to and takes from its environment. Thus, there is a range of interdependent organisms. In the words of Baskin, ‘It is this lavish array of organisms that we call “biodiversity”, an intricately linked web of living things whose activities work in concert to make the earth a uniquely habitable planet.’11 Just as a body depends upon division of labour among cells, so an ecosystem depends upon division of labour across a diversity of organisms. Without biodiversity services, there would be no ecosystem and probably no life. Thus, biodiversity studies look at entire ecosystems. The term, ‘balance of nature’ was and still is frequently used. We now know that this ‘balance’ involves a tighter web of relationships than previously imagined.
The major emphasis of biodiversity studies has been saving endangered species. It came to be realised that trying to save species, one at a time, was literally a dead end approach. Species exist in ecosystems and depend upon them. Thus, endangered species could not be saved without also preserving their ecological support systems. Moreover, required species services necessitate whole populations of species that provide them, not just a few individuals. Saving only a token number of individuals, like stamps in an album, will not sufficiently provide the services. Seriously reducing these populations, or their total loss, can only have far-reaching, often dire, consequences.
As information about biodiversity services accumulated, and the value of species in ecosystems became evident, some have become aware that such services and the interdependence of species derived from those services, speak beyond the immediate needs of ecosystems to the origin of ecosystems and ecology and even of life itself. However, very few have made such connections; the immediate conservation problem has been the primary focus. However, when the vital nature of collective interdependent ecological relationships and services is considered, they appear designed; further suggesting that they must have ‘originated rapidly because complete … ecosystems are necessary for the survival of living things [emphasis added]’.12
In recent decades, especially since focused efforts to save endangered species and environments began, biodiversity information has been accumulating. Unfortunately, much of the information has come from ecosystem damage and destruction.13 That is, after species become extinct or rare, it has been easier to see, in some degree, what their ecological roles had been because of the ‘ecological holes’ their absence produced. Nevertheless, regardless of how information was gathered, it is now known that ecosystem members necessarily interact with each other to provide mutually supporting services. Ecological interdependence is tight enough that, according to Peter Raven of the Missouri Botanical Garden, when a plant is exterminated, often as many as ten to thirty other creatures follow it into extinction.14
Ecological interdependence
Every living organism, without exception, has needs that must be met. Some needs may be met by internal mechanisms, but many others are met from their environment, frequently from other organisms. There is an interdependence among creatures that is only now becoming apparent.
A number of authors have listed ecological services provided by different interacting organisms. However, it has usually been possible to add to the lists, so it is certain that no one has a full grasp of the range and details of ecological services. Nevertheless, a short list might suggest just how pervasive ecological services and needs for them are. Most listed services are broad categories under which are many detailed interactions and a vast array of different species.

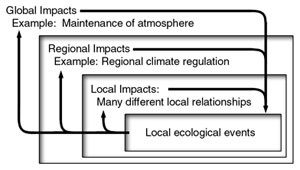
The list includes: detoxification of air and water; alleviation of floods and droughts; renewal of soil and soil fertility; pollination services; dispersal of seeds and other dissemules; provision of nutrients; maintenance of biodiversity; protection against UV radiation; stabilisation and moderation of temperature—locally in what may be termed ‘micro-climates’, regionally, and even globally; rainfall regulation; regulation of evapotranspiration; pest control; recycling of essential nutrient minerals; capture, alteration and provision of energy; physical support; water supply services; provision and development of new habitats; expansion and use of potential niches; erosion control; provision of nest sites and materials; camouflage materials and services; albedo (reflectance) reduction; protective cover; and so on. Each organism receives and gives as illustrated in Figure 2, focusing on a single tree. Some of the services are indispensable for survival of dependent species. Most of the services are beneficial to all concerned and, certainly not harmful.
Careful analysis of ecology reveals that, while ecological services may appear to function locally, (see Figure 3), they may also have far-reaching and reciprocating influences. The extent of ecological services may not be immediately apparent.
Many services are held to be necessary for the well being and survival of individuals, and by extension, entire species populations, not to mention other dependent species. Each service may vary in detail from one ecosystem to another, depending upon where and by whom it is provided. Specific species are not always essential for the provision of specific services. That is, when more than one species in an ecosystem is able to provide the same service, if one of those species is missing or declines, survival of the dependent species may still be assured. However, this is not to say that such ‘redundant’ species are expendable, as some have asserted. Since most, if not all species appear to provide more than one service, and may not be totally redundant, each species may still be essential for the survival of other species. Moreover, redundancy provides a back up for times of difficulty when one of the providers may be disabled.
Several specific examples of ecological interdependency may be helpful. Consider soil. Many undoubtedly think of soil as a relatively inert environment. Yet, in that same soil, there is frenetic life-supporting activity involving myriads of microbes. Nevertheless, soil organisms and their varied functions remain largely unknown and we abuse soils, ignorantly perhaps, to our own hurt.
Soil fertility services are provided by an enormous number of species of bacteria, annelid worms and nematodes, fungi, insects, mites, and millipedes, crustacea, algae, protozoa and more,15 each with different tasks, like workers on an assembly line.16 The populations of these species are huge. Each set of workers makes use of its specific abilities to do precisely what needs to be done to provide essential products for the next step in the process that is managed by a different group of microbes. ‘Ecochemical pathways’ thread across the soil community and even beyond into the supraterranean ecosystem, to become a bulwark for life itself.
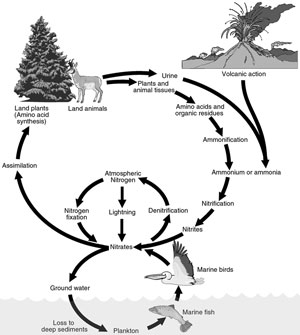
Consider a specific example: the nitrogen cycle.17 It is one among many different soil maintaining processes. Generations of students have learned details of this cycle, and it is an outstanding example of the bacterial assembly line. Atmospheric nitrogen is reduced to ammonia by nitrogen-fixing bacteria, such as Rhizobium in root nodules of legumes that actually grow in response to the bacteria (How did that happen?), by associations of bacteria in roots of other plants, such as alders and Casuarina, in marine algae, in lichens, and even in shipworms, and by Azotobacter, and by several species of cyanobacteria, some of which are also found in association with plant roots. Ammonia is also provided by ammonification or hydrolysis of protein wastes by all organisms.
Other soil bacteria occupy different places along the assembly line, participating in nitrification, or the conversion of ammonia to nitrites and nitrates. Nitrosomonas in soil, and Nitrosococcus in seawater, convert ammonia to nitrites. Nitrites are changed to nitrates by Nitrobacter in the soil or by Nitrococcus in the sea.
Plants assimilate ammonia and nitrates, and convert them into various essential nitrogenous compounds like proteins and nucleic acids. These are then passed on directly to herbivorous animals and indirectly to carnivorous animals, and so on.
Finally, denitrifying bacteria, like Pseudomonas denitrificans, reduce nitrates to nitrous oxide and atmospheric nitrogen. The cycle has gone full circle (see Figure 4).
The important point for our purpose here is that many bacterial species are involved, but no single organism is able to do everything. Each step is performed by different organisms that must ‘cooperate’ with each other. Each organism has a different set of enzymes that enables it to do its specific job in the array of tasks that must be completed. Each one is provided for, but at the same time, provides for others. If any part of the process were missing, all would be hurt.
Abiogenic nitrates are also formed in thunderstorms. Robert Leo Smith18 estimates that thunderstorms and cosmic radiation account for only 10 % of nitrogen fixation. This is also an inconsistent source. Moreover, many creationists believe violent storms were foreign to original conditions. Thus, it seems that there could be neither plants nor animals, nor life at all, without microbes capturing, processing and recycling nitrogen, for neither plants nor animals fix nitrogen.
Other nutrient cycles likewise require services of bacteria and fungi. Without fungal wood-rotting services,19 for example, carbon would accumulate as cellulose and lignin. Without decomposers, cellulose would endure indefinitely and gradually reduce atmospheric levels of CO2, which, in turn, would reduce photosynthetic activity in plants. Eventually, all living systems would be impacted, if indeed, they were ever able to function at all.
Another example of plant dependence—one among many—is the relationship plants have with mycorrhizal fungi. It has been noted that nearly all plants either have or could have mycorrhizae.20 Mycorrhizae are in intimate contact with root cells and extend into the soil to become conduits for nutrients that plants with the same sized root systems would otherwise not be able to access. They expand the absorptive surface for nutrient procurement. Mycorrhizae and their host plants also interact in a variety of other ways to benefit plants, including disease suppression.20 F.L. Pfleger, with R.G. Linderman, research leader of the Horticultural Crops Research Laboratory, of the U.S. Department Agriculture, says that plants in ecosystems are highly dependent on their mycorrhizae for survival.20 They suggest that they evolved together. Might we say, created together? They appear to be that necessary.
We speak of plants as being autotrophic, or ‘self feeders’. In terms of capturing energy, this appears to be appropriate, but this also seems to be the only place where plants are really independent. Plants may vary in the details of their dependencies, but in general, plants are as dependent as any animal on the availability of their mineral nutrients, carbon included (although it is not organic carbon).
Plants use the products of decomposition, and are particularly dependent on other organisms with respect to atmospheric nutrients—carbon dioxide and nitrogen.21 They, in turn, recycle oxygen. While other soil nutrients may be made available by erosion and leaching, they are also subject to recycling by microbial decomposers. Erosion alone, like thunderstorms for nitrogen, is an unreliable source now, and would have been an unlikely source under early climatic conditions which would have minimised erosion.
These are a few examples of interdependence in nature. Numerous other examples could be given involving pollinators, seed dispersers, relationships between plants and ants or other insects, and so on. As nature is now, these numerous and varied relationships are essential for survival of many species. The current necessity of ecology suggests that ecology has always been necessary, although the negative relationships we find today would have been lacking in pre-Fall ecology.
Biodiversity research
With the rise of interest in biodiversity, a few scientists have endeavoured to quantify the nature and importance of biodiversity services. Their work has considered the relationship between biodiversity and such phenomena as ecosystem productivity, soil fertility, community stability, sustainability and resiliency. An excellent review has been written by David Tilman.22 The following discussion is based on his review.
As biodiversity increases, so does community productivity, but at a declining rate, until the ecosystem reaches saturation and productivity levels off. At this point, even if species richness continues to increase, productivity does not.
Click here for larger view
There are two possible explanations for this. Firstly, with greater biodiversity, there is greater probability of having more highly productive species present, particularly in relation to season (e.g. some plant species grow best in summer, others in winter). Secondly, because of the different requirements of different species, greater biodiversity allows fuller exploitation of available resources. Consequently, the area is used more efficiently. However, the rate of increase in productivity declines as biodiversity increases. Once saturation is reached, the impact of individuals upon others in the community, limits individual productivity to the point that overall community productivity remains at plateau level despite the increasing diversity of species.
As species richness increases, the variance in productivity among different species also increases, but the collective stability of all species together, the complete ecosystem, increases. That is, total ecosystem variance declines with increasing biodiversity.
All ecosystems are subject to disturbance. However, the more diverse an ecosystem is, the more resistant to disturbance it appears to be—with the potential for a drop in productivity being notably less than for ecosystems with fewer species. Evidently, this is due to the presence of some plants that are less stressed by specific disturbing conditions than others. Less diverse ecosystems, on the other hand, will be less likely to have such resistant species present. Under another kind of disturbance, however, ecological roles may be reversed so that formerly negatively impacted plant species may be able to function and cover for the other, now less productive plants. That is, one plant’s disturbance may be another plant’s opportunity.
There is ambiguous evidence that more diverse ecosystems may be more resilient or better able to recover after disturbance. Some studies support the idea, but others appear to neither support nor negate it. Consequently, the relationship between resiliency and biodiversity is still unclear.
In each ecosystem, resources are limited. However, more highly diverse ecosystems are able to use resources more efficiently. This efficiency leaves fewer soil nutrients available for leaching into ground water to be lost. Experimental work reveals that different species have different effects on nutrient cycling. Thus, soils of more diverse ecosystems tend to be more fertile. It was suggested that when there is greater above ground diversity, there may also exist higher soil decomposer diversity. The above ground increased biodiversity produces a greater variety of substances which can support a greater variety of decomposers.
Tilman sums up the review with this statement: ‘Dependence on biodiversity is no magical effect, but rather reflects the increased functional roles that are possible in more diverse ecosystems.’ The important point for our purpose here is that the proposed importance of biodiversity services in ecosystems stands up under close scientific scrutiny.
Redundant services
Some services are offered by more than one species. Thus, some species appear redundant in terms of their services.
In 1981, pioneers in biodiversity studies, P.R. and A.H. Ehrlich, wrote about the importance of different species to ecosystems. 23 They likened species to rivets in an aeroplane and pointed out that, like aeroplanes, ecosystems tend to have redundant subsystems and other design features that allow functioning to continue even after a certain amount of abuse. This idea became known as the ‘rivet popper’ hypothesis. It was understood to emphasise the importance of each species and, as a consequence, people lost sight of the idea of redundancy.
In 1992, and again in 1995, Australian ecologist, Brian Walker,24 saw that not all species in ecosystems are of equal functional importance. He observed that different species were in different functional groups in which, evidently, some species could be eliminated without obviously disturbing the functioning of the ecosystem, since more than one species offered the same service. This became known as the ‘redundancy’ hypothesis.
In a paper they published together in Bioscience,25 both Paul Ehrlich and Brian Walker together attempted to correct certain misunderstandings. The Ehrlichs had previously noted redundancy also, but had emphasised general ignorance regarding which species might be redundant and possibly expendable. While an ecosystem may be able to absorb abuse without obvious loss of function, Ehrlich and Walker point out the importance of redundancy in maintaining resiliency. Different species are able to offer the same service, but may do it under different circumstances so that they actually replace each other when necessary. The immediate value of a species, as pointed out by Tilman, may change, depending upon current conditions. Ehrlich and Walker also noted that conservation emphasis should be toward saving species whose ecosystem functions were observed to not be redundant. Nevertheless, regardless of redundancy, as they said it, ‘we force species and populations to extinction at our own peril’. Peter Raven asserts that ‘even in simple ecosystems, biologists cannot tell as yet which species might prove superfluous’.26
Superficially, it might appear that the presence of redundant systems is wasteful and, consequently, a failure of the ‘design hypothesis’ for the origin of ecology and ecosystems. Would redundancy, under ideal conditions, have been necessary at all? How could its origin be explained in a design model? Is redundancy a challenge or is it explainable as part of the design hypothesis?
While redundancy may seem out of harmony with the ideal, it undoubtedly had a place in the original scheme of things. Redundancy is found everywhere in nature, after all, not just in ecology. So, how can ecological services redundancy be explained within an ecological design model?
Firstly, some services are needed in amounts that cannot be provided by only one species. Redundancy would be necessary. Indeed, some services are provided by a great many different species—e.g. photosynthetic products.
Secondly, original environments may have lacked extreme conditions found today, but they may not have been uniform either. With environmental differences, certain species would have been more suited to some situations than others. Thus, different species for different environments, might have offered redundant, but not overlapping services. When nature began to suffer and breakdown, redundant services offered by species that formerly occupied different environments may have been forced together.
Thirdly, understanding redundancy today involves a distinction between what we deem a species now and what the Creation account refers to as a ‘kind’. They should not be considered identical and indeed, most creationists do not equate them. Thus, numerous species are thought to have arisen from each created kind. Surely many of these would provide similar, if not redundant services.
Original ecology
That ecology was created is suggested in the Creation account, where food procuring and reproductive relationships are described.27 Without doubt, original ecology must have been different from what we find today. Creationists consider that climate was mild, there was no rain28 or violent storms, and death29 was absent. Without death, for example, what would regulate populations? Population regulation could have been due to ecological mechanisms that no longer exist or function. Gene pools must have been large, given the diversity of fossils, and what we know would have happened to gene pools with massive loss of life. Thus original ecosystems appear to have had high biodiversity, indicating that original ecology may have been vastly more complex and different than ecology now.
The original ecology may be seen as dynamic, with producers, consumers and decomposers —for did not man and animals eat plants, and were there not wastes to recycle? Did leaves fall from trees or was the abscission layer in the petiole an after-thought? Not all fruit was eaten surely, and there are inedible portions of fruits.
Early life reproduced and expanded to eventually fill earth, as instructed. In this process, they would have worked together, alternately establishing and filling niches. This process of expansion would, in certain ways, have been a successional process, although probably different from succession today. Wherever you turn in the Creation account, you bump into ecology.
Sin and, particularly the Flood, introduced conditions that would have been challenging to nature. Indeed, Romans chapter 8 tells of suffering and corruption that came to both man and nature. After the Flood, life had to be established anew on Earth's surface. How could this happen?
During the Flood, air breathing terrestrial animals died (Genesis 7:22), but many other organisms—marine and aquatic animals, microbes, and plants must have survived, although many of these also perished. Not all land was flooded for the same length of time, nor did the Flood end everywhere at the same time. Highlands and mountains were pushed up, it appears, and ocean depths opened. Waters poured off the land. Animals in the ark were freed to reproduce and spread out.
After the Flood, the stage was set for a successional return to a semblance of communities and ecosystems. These would not be exactly like those before the Flood, however, for there had been many changes due to extinctions and gene loss, mutations, and topographical and climatic changes. In isolation, in biological islands at first, many new species formed. New and also negative relationships formed. Nevertheless, essential ecological services must have been available.
Negative relationships in nature have suggested to some that ecology is a product of degeneration. For them, there was but little ecology at first. They picture ecology as coming from the breakdown of nature. The introduction of death30 unquestionably was a new dynamic. It resulted in such degenerate phenomena as scavengery and carrion eating, predation with carnivory, and parasitism with disease.
However, these may possibly be seen as resulting from a breakdown of relationships that had originally been benign and even mutually beneficial. Moreover, extinction of species and loss of genes would have contributed to the development of negative relationships by severely reducing gene pools and restricting ecosystems. With loss of resources and community cohesion, survivors would have turned to other less than optimal resources they were never intended to use or need.
Negative relationships, however, should not be used to question that there was a created ecology, any more than one would question the creation of fully functional metabolic pathways because there are now genetic diseases and defective enzymes.
Biodiversity, redundancy and design
The idea of design in nature is an old one. Undoubtedly, readers of this journal are familiar with the name and contributions of William Paley (1743–1805). He wrote of natural design and natural theology and thought such were evidences for God.
Darwin read Paley; (his books were required reading at the university) and was evidently influenced, but not persuaded by him. Some of Darwin’s writings, consequently, were specific challenges to Paley’s ideas. Under the influence of Darwinism, the impact of Paley diminished, but the power of his argument is still felt today. Why, for example, did Richard Dawkins write the Blind Watchmaker?31 Why did he find it necessary to contest the writing of an individual who lived close to 200 years earlier, if his ideas did not still have power? Dawkins attempted to show that what appears to be design in nature is really the product of natural processes. Having rejected other possibilities, what else could he do? This goes to show to what lengths some will go to side step the most obvious and compelling evidence to the contrary.
Dawkins appears to be locked into a philosophy that leaves no other choice but to explain life and nature as a product of natural events. However, while he may be comfortable with this position, other scientists are not. Lewis Thomas well stated the dilemma when he wrote:
‘I cannot make my peace with the randomness doctrine; I cannot abide the notion of purposelessness and blind chance in nature. And yet I do not know what to put in its place for the quieting of my mind. It is absurd to say that a place like this place is absurd, when it contains, in front of our eyes, so many billions of different forms of life, each one in its way absolutely perfect, all linked together to form what would surely seem to an outsider a huge spherical organism.’ 32
What is the answer for Lewis Thomas and other scientists like him? Behe appears to have an answer. He finds that complex biochemical pathways in cells suggest design.
Some have found it possible to accept design as the explanation for complexity at the biochemical level, but then reject creation of different species. For them, evolution is the accepted explanation for the complexity of living organisms. However, ecology, with its ecochemical and ecophysical pathways, and relationships that are too complex and too essential to easily be accounted for by gradual development, negative relationships notwithstanding, similarly challenges the idea that evolution explains diversity of organisms.
If evolution of a cell or organism is compared to a computer accidentally assembling as a result of an explosion in an electronics warehouse (I have heard this comparison), the evolutionary development of ecosystems would have to be compared to the world-wide web, together with all its necessary and integrated computers, wiring, software and wealth of information, resulting from a series of catastrophes occurring independently in many electronics warehouses. No one doubts that computers and the worldwide web were designed, but then, many doubt that natural systems, including ecosystems, which are even more complex, were designed.
When the Ehrlichs proposed the ‘rivet popper’ hypothesis, they noted that ‘Ecosystems … tend to have redundant subsystems and other design features that permit them to continue functioning after absorbing a certain amount of abuse [emphasis supplied].’ There it is again, ‘design’!
Baskin responded to the Ehrlichs:
‘One problem with likening species to rivets, however, is that it implies there is a design, that species are deliberately placed where they are needed. Yet our current understanding of how communities are assembled says that species move in and evolve as opportunities present themselves; in other words, when they can, not when they are needed. The role of species may also change from one community to another or with shifting conditions.’ 8
Baskin is correct, of course, in her description of community dynamics, but she is too narrow in her assessment of the word, ‘design’. Communities appear to have a dynamic design and are able to develop and enlarge over time—a process known as succession—as preceding species provide necessary services for succeeding species. But note, ecological services, provided by biodiversity, must already be available somewhere in order for ecological succession to occur.
Amazingly, even when the development of a community must begin on bare rock, as after a volcanic eruption, possibly in the middle of the sea, or after the Flood, when pioneer plants arrived on the scene, microbial decomposers were already present. Symbiotic associations are needed at each stage of development. The design not only accounts for the end result, but also for how it developed over time, often a long period of time.
The idea of design does not require that species in a community have a specific place. In fact, specific services often seem more necessary than specific species. Nevertheless, it is difficult to get away from the idea of integrated services being required for ecosystems to exist now, as well as throughout the period of life’s existence on earth.
Undoubtedly, the Ehrlichs did not intend to suggest a designer when they used the word ‘design’ in their ‘rivet popper’ paper, but no other word adequately conveys the idea. If something looks designed, however, it most probably was designed. If something looks designed, why not think of a designer as a possible explanation? Indeed, the functional design of ecosystems becomes increasingly obvious the more they are studied.
Conclusion
Since ecology is built upon incredible multi-species complexity, trying to explain the origin of the wonderfully integrated diversity of life by chance events painfully stretches one’s credulity. Everywhere we look we can find evidence of life having been designed by a Creator, even at the ecological level. This leads to two important conclusions:
- the necessity of ecology in the operation of ecosystems today suggests that ecology has always been needed;
- ecological services had to have been provided quickly in order for original ecosystems to operate. While this may not specifically demand a six-day Creation, it is in harmony with it.
The incredible interdependence between species—systems of living things supporting each other—is exactly what we would expect to find from a Creator who ‘makes grass grow for the cattle,’ and brings forth ‘food from the earth’ (Psalm 104:14), and ‘who gives food to every creature’ (Psalm 136:25). It is He who said ‘Give, and it shall be given unto you’ (Luke 6:38), and, ‘Freely ye have received, freely give’ (Matthew10:18). Do not these contain the essence of ecology—giving and receiving—and the modus operandi of heaven? Biodiversity is indeed a powerful testimony about the Creator that reminds us of the truth of Romans 1:20. ‘For since the creation of the world, God’s invisible qualities—his eternal power and divine nature—have been clearly seen, being understood from what has been made … ’
References
- Psalm 104:24, 27, The Holy Bible, New Century Version, Word Books, 1990. Return to text.
- Smith, R.L., Elements of Ecology, Harper Collins, New York, p. 3, 1992. Return to text.
- Smith, ref. 2, p. G3. Return to text.
- Behe, M.J., Darwin’s Black Box, Touchstone, Simon & Schuster, New York, NY, 1996. Return to text.
- Javor, G., George Javor, Biochemist; in: Ashton, J.F. (ed.), In Six Days: Why 50 Scientists Choose To Believe In Creation, New Holland Publishers (Australia) Pty Ltd., Sydney, pp. 123, 124, 1999. Return to text.
- Reported in the November, 1986 edition of Smithsonian Magazine. Return to text.
- Tilman, D.; in: Daily, G.C. (ed.), Nature’s Services: Societal Dependence On Natural Ecosystems, Island Press, Washington, D.C., p. 94, 1997. Return to text.
- Baskin, Y., The Work of Nature: How the Diversity of Life Sustains Us, Island Press, Washington, D.C., p. 6, 1997. Return to text.
- Daily, G.C. (ed.), Nature’s Services: Societal Dependence on Natural Ecosystems, Island Press, Washington, D.C., 1997. Return to text.
- Calculations by Hughes, J.B., Daily, G.C. and Ehrlich, P.R., as cited in Science News 152(17):260, 1997. Return to text.
- Baskin, ref. 8, p. 3. Return to text.
- Sunday Herald Sun, Melbourne, 6 June 1999. Return to text.
- Daily, G.C., Introduction: what are ecosystem services? in: Daily, ref. 9, p. 5. Return to text.
- Raven, P.H., Ethics and attitudes; in: Simmons J., et al. (eds), Conservation of Threatened Plants Plenum Publishing, New York pp.155–181, 1976; cited by Baskin, ref. 8, pp. 36, 37. Return to text.
- Daily, ref. 9, p. 116. Return to text.
- Baskin, ref. 8, p. 110. Return to text.
- Much of the information here has been taken from Ricklefs, R.E., Ecology, W.H. Freeman and Company, New York, pp. 220–226, 1990. Return to text.
- Smith, R.L., Ecology and Field Biology, 5th ed., Harper Collins, New York, 1996. Return to text.
- Ricklefs, ref. 17, p. 225. Return to text.
- Pfleger, F.L. and Linderman, R.G., General summary from Mycorrhizae and plant health, 1998, from the Horticultural Crops Research Laboratory in which Dr. Linderman is research leader; <URL: http://www.ars-grin.gov/ars/Corvallis?hcrl/hcrl.htm>. Return to text.
- Aside from the small amount produced by thunderstorms. Return to text.
- Tilman, D., Biodiversity and ecosystem functioning; in: Daily, G.C. (ed.), Nature’s Services: Societal Dependence On Natural Ecosystems. Island Press, Washington, D.C., pp. 93–112, 1997. Return to text.
- Ehrlich, P. and Ehrlich A., Extinction: The Causes and Consequences of the Disappearance of Species, Random House, New York, 1981; as cited in Ehrlich, P. and Walker, B., Rivets and redundancy, Bioscience 48(5):387, 1998. Return to text.
- Walker, B.H., Biodiversity and ecological redundancy, Conservation Biology 6:18–23, 1992; and Walker, B.H., Conserving biological diversity through ecosystem resilience, Conservation Biology 9:1–7, 1995; as cited in Ehrlich, P., and Walker, B., Rivets and redundancy, Bioscience 48(5):387, 1998. Return to text.
- Ehrlich and Walker, ref. 24. Return to text.
- Cited by Tangley, L., in US News & World Report 123(16):68, 1997. Return to text.
- Genesis 1:22, 27, 29 and 30. Return to text.
- The first appearance of the rainbow after the Flood would support the no rain hypothesis. Return to text.
- A distinction could be made between cell death and organismic death. Return to text.
- I make a distinction between cell death and organismic death. Originally there was cell and tissue death, for animals ate from plants and one might infer decomposer too, to break down wastes. Even those decomposer cells may have died in the exercise of their functions. Return to text.
- Dawkins, R., The Blind Watchmaker: Why the Evidence of Evolution Reveals a Universe without Design, W.W. Norton & Co, New York, 1990. Return to text.
- Thomas, L., On the uncertainty of science, Harvard Magazine 83(1):19–22, 1980; as quoted by Roth, A., Origins: Linking Science and Scripture, Review and Herald Publishing Association, Hagerstown, MD, p. 333, 1998. Return to text.
Readers’ comments
Comments are automatically closed 14 days after publication.