

Journal of Creation 18(2):114–120, August 2004
Browse our latest digital issue Subscribe
Human and dinosaur fossil footprints in the Upper Cretaceous of North America?
Abstract
The Upper Cretaceous Dunvegan Formation of British Columbia has yielded some of the best dinosaur footprints in Canada. The area near Tumbler Ridge is one of the few locations in the world with dinosaur footprints and dinosaur bones on the same bedding plane. Human-like footprints were recently discovered in the area, too, but on closer examination they seem to be metatarsal dinosaur footprints. The discoverers of these ichnofossils also found some of the best dinosaur trackways in Canada, described for the first time in this paper.
Ichnology (Greek ίχνος ichnos = footprint) is the science that deals with the tracks, trails, burrows and other traces left by living animals, and in many respects could be compared to forensic science. While some paleontologists see the study of animal tracks as a ‘lunatic fringe of paleontology’1 pursued by bands of enthusiastic amateur track hunters, most paleontologists recognize ichnology as a worthwhile endeavour and a valid branch of paleontology.

Interestingly, people who study tracks of present-day animals are generally described as ‘trackers’; however, when the study becomes scientific the name turns into ‘biophysicist’. There is rarely a problem in finding the ‘authors’ of the tracks of present-day animals, but when it comes to the fossil tracks of extinct animals, the problem is much more difficult. There are millions of extinct-animal traces and tracks (ichnites), but there are extremely few skeletal remains (in the case of vertebrate ‘authors’) on the track-bearing layers. And often the creatures, the bones of which are most common in a given geological setting, have left few or no tracks (as in the case of ceratopsids2). Thus the animals that left the fossil tracks are usually unknown or guessed at.
This lack of matching between extinct animals and fossil tracks has led to a multiplication of fossil names. Thus, trace fossils, like body fossils, are given individual Latin names using the Linnaean binomial system, even though the trace in itself does not represent a unique organism. So names like Baropezia fontis emerged designating an unknown ‘heavy-footed (broad-toed) creature from the Fountain Formation’ (Carboniferous of Colorado).3
Another example is Laoporus (‘stone tracks’) from the Coconino Sandstone in the Grand Canyon. Most paleontologists attribute Laoporus to amphibians, many of the features of which resemble the tracks that salamanders, manoeuvring in shallow water, produced during experiments. Despite the difficulties explaining away this evidence for a wet environment, many geologists still interpret the Coconino Sandstone as a desert formation!4 Most creationists believe the geomorphology of the Coconino Sandstone and other formations found in the Grand Canyon, with their perfectly conformable flat contacts, massive areal extents and limited thickness (in relation to its extent, the Coconino is thinner than this page), represents a continental-scale Flood deposit, i.e. Noah’s Flood. Despite the united efforts to dismiss the subaqueous origin of the Laoporus, one cannot forget the case of the formations in the north-eastern Bighorn Basin in Wyoming. Interestingly, these were described as marine in origin until dinosaur tracks were recently discovered in them!5,6
As would be expected, the most dynamic area of ichnology deals with dinosaurs, and the number of paleontological names, or ichnotaxa, is rapidly catching up with the already bushy taxonomy of dinosaurs.7 Doubling the number of taxa on the grounds of footprints, which most probably belong to already-known taxa, is unlikely to make our understanding of dinosaurs simpler and more accurate.
The Dunvegan Formation
The Dunvegan Formation (DF) is interpreted as a deltaic complex of Cenomanian age, advancing into the sea about 400 km from NW to SE (from British Columbia to Alberta), over about 2 million years.8 The formation is described as prograde, based on a coarsening-upwards grading from clays to sands to gravels, interpreted as the result of the changing relative deposition location from off-shore to near-shore. According to some authors, the DF covers a much larger area (about 300,000 km2), extending as far as the Northwest Territories.9 Lithologically, it consists of interbedded mudstone, sandstone and conglomerate. Coal, either as lenses or irregular bodies, is frequent, but in amounts significantly smaller than the Lower Cretaceous coal deposits which are mined in the area.
A creationist interpretation of this formation would be a primarily depositional environment under fluctuating water levels during the Flood, as discussed later.
The footprints

The first theropod footprints (ichnofossils) in the DF were reported in 1975 from the Pine River area in British Columbia.10 In the summer of 2000, following flooding of Flatbed Creek (near Tumbler Ridge) that cleared a rock face, two local boys discovered dinosaur footprints, which were later assigned to ankylosaurs (ichnotaxa Tetrapodosaurus borealis), the first ever found. The footprints are grouped in a trackway, which is the first in situ trackway found in B.C., as all previous ichnites have been found in blocks of rock dislocated from their original position. In addition, the Flatbed Creek site yielded a few dinosaur bones (attributed to ankylosaurs, but there was insufficient data for a solid identification).
Subsequently, footprints were found in the left bank of the nearby Wolverine Creek, this time belonging to ornithopods. Figure 1 shows the ichnofossil sites in the Tumbler Ridge area.
The Wolverine Creek site
Recently, creationists Fred and Ruth Walkley, and Bruce and Joan Zimmerman from Tumbler Ridge discovered two new track sites in this area. They noticed a strong resemblance of some of the ichnites to human footprints. To their credit, they decided not to reveal the site until a professional geologist with a Christian worldview could see it first. Following their invitation, I visited the site in early October 2003.
The presumed human prints were located in the left bank of the Wolverine Creek, about a mile upstream from the Wolverine Bridge (figure 2). The site was exposed after a flood in the spring of 2001 washed away overlying material. It has a triangular shape (figure 3), bordered by the river to the S–SW, a vertical outcrop to the N–NE and a rubble heap to the E–SE. The outcrop reveals interbedded mudstones, sandstones, marls and shales (figure 2). The shales in the upper part are black and contain frequent ferruginous nodules. Also a white film (possibly montmorillonite) covers the outcrop in a few locations after seeping out of the strata.

The secular paleontologists working in the area had visited the site before us and concluded there were no ichnites there. I disagree with their conclusion and believe they simply overlooked them because they were expecting to see footprints complete with toe marks (digitigrade ichnites—like prints made by cats and dogs).
The ichnites appear on a highly irregular bedding plane of a dark grey mudstone (figure 4). I investigated seven features (labelled WOC 01 to 07) of which five have strong resemblances to human footprints, but are much larger. No trackway was found, not even two consecutive steps that could allow any estimation of pace, stride, gait, etc.
One of the features (WOC 05) has one characteristic that rules out its being an ichnite: the bottom plane is horizontal while the bedding plane dips 20° to the SW (220°). This strongly suggests a postdepositional feature (possibly erosional). WOC 03 was half under water and WOC 04 completely covered by water, so the only ichnites that allowed the full set of measurements were WOC 02 and 06.
WOC 02

The general outline of WOC 02 (figure 5) is similar to a human right footprint. Though not evident, an arch can be inferred, which is the only reason to consider it a ‘right footprint’. There is a clear secondary, wider contour of the frontal and upper right side of the print which suggests originally soft, wet sediment. No toeprints are present. The maximum depth of the print is 30 mm towards the upper right side, gradually decreasing to close to 0 mm at the heel. This suggests the walking manner was on the whole sole of the foot, like bears or people (plantigrade or quasi-plantigrade walking manner) rather than walking on the toes (digitigrade walking manner). Most ichnites found in the area have been digitigrade with 3 or 4 toe prints, tri- or tetradactile.
Kuban has pointed out that bipedal dinosaurs, which normally walk digitigrade, may have occasionally walked in a plantigrade manner, especially when on soft, fairly deep sediments.11 The detailed bedding plane morphology of the ichnite-bearing layer (especially its ruggedness) suggests the sediment was soft and prone to slumping. Footprints would have been very poorly preserved. I suspect some postdepositional process, such as loadcasting (where sandy material protrudes into underlying finer material), further distorted the original prints.

The bottom plane of this print is parallel to the bedding, and its size well exceeds any normal human footprint. The bearing of the print is due west (270°).
WOC 06
The print of WOC 06 (figure 6) is less clear and quite symmetric, which makes any identification as ‘left’ or ‘right’ foot impossible. The ball section is broader than that of WOC 02. The maximum depth—located in the mid section of the print—is 46 mm, gradually decreasing toward the heel, which is actually very difficult to outline. No toeprints are present. The print’s bottom plane is parallel to the bedding, and again the print is larger than any normal human footprint. The bearing of the print is south-west (221°). There is no visible secondary, outer contour as in the case of WOC 02.
Comparison
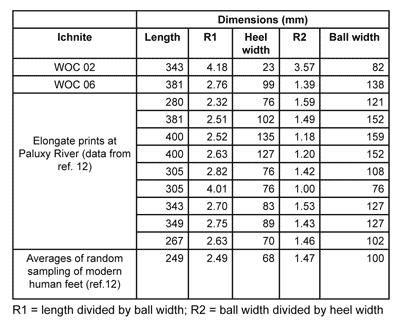
WOC 06 is longer, wider and deeper than WOC 02, and the angle between the orientations of the two prints is 49°. The anatomic ratio differences are presented in table 1.
There are marked differences between R1 and R2 for WOC 02 and 06. And given the different bearing and general morphology, the data indicates different authors. I have introduced comparative data from the Paluxy River ichnites, which shows similar morphology and is the only dataset providing acceptable morphometric information. However, from what I could find in the literature, the Paluxy River ichnites are probably not human, but dinosaurian. Consequently, the differences are rather irrelevant for the above comparison.
The Paluxy River trackways

The Paluxy River site, as well as the nearby Dinosaur Valley State Park’s ‘Taylor Site’ and ‘Shelf Site’ are well known and have triggered a hot debate between some creationists and evolutionists.12 Some creationists have suggested that the elongated, indistinct depressions alongside clear dinosaur tracks are human footprints, based on morphological arguments (i.e. the resemblance to human footprints). The anatomical ratios (see table 1), when compared to modern humans, are not at all convincing. In fact all ichnites are much larger than the modern human foot.

Most problematic of all, the pace is bigger than that of the largest modern humans—over 1 m.13 Running cannot be invoked because of the soft, muddy sediment in which the prints were produced. It is virtually impossible for a human to run with a pace of over one metre on such a substrate! Furthermore, the indistinct, elongated tracks are a minority compared with the clear tridactyl elongated tracks which represent most of the individual tracks, within any trackway (figure 7).
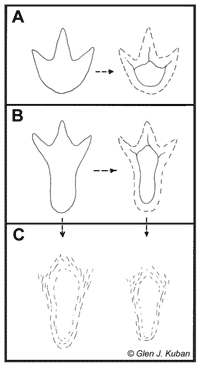
I find Kuban’s interpretation of the indistinct, elongated tracks from Paluxy River (PR) as metatarsal dinosaur tracks resulting from mud collapse very plausible.14 There are many locations around the world displaying elongated tridactyl dinosaur prints.15 In most of these cases they alternate with normal digitigrade tridactyl ones (figure 8). It looks like the animals were walking on soft sediment and every now and then bent their knees more than usual so that the metatarsals reached the ground, increasing the supporting surface. Some authors interpret metatarsal dinosaur tracks as indicating a crouching behaviour.16 Because of the soft, waterlogged substrate, soon after the animals passed, the contour of their feet would start slumping and collapsing until, in some cases, the impression of the digits was completely lost and only an elongated hollow, wider at one end, would remain. After diagenesis, when such hollows would become exposed to weathering, their contours would get even more fuzzy and further lose the original features. Figure 9 illustrates such an interpretation.
Personally, I lean towards interpreting WOC 02 and 06 as metatarsal dinosaurian footprints, too, although I would not completely rule out the possibility of them being human. However, without a sequence of at least three consecutive prints, I would not even consider trying to build such a case.
The second site
The ichnofossil discoveries in the Tumbler Ridge area have unfortunately attracted some individuals with a definitely different philosophy of track hunting, namely ‘take ’em with you’. Since my visit in the area, a theropod footprint at one of the sites has disappeared, having been pried out from the rock layer—one of the very few in situ dinosaur footprints in the whole of Canada! Because of this I have chosen not to reveal the location of Ruth Walkley’s discovery in figure 10. Unfortunately, this is not in situ either. This splendid trackway is located on a large slab of mudstone and composed of four clear footprints and two clear handprints. Adding to the value of this trackway is the fact that the entire slab surface is ripple marked; a very clear and rather precise paleoenvironmental marker which will be discussed later.

The general appearance of the footprints suggests a theropod, but the many-fingered handprints seem to rule out such a possibility. Theropods were mainly bipedal and their hands had a different morphology. There is another characteristic that raises doubts as to the theropod origin of this trackway: although the length of digit III is greater than that of digits II and IV, the difference in length is smaller (only 2 cm) than in usual theropods. The posterior end of the footprints is almost linear (figure 11) and not v-shaped as with most of the theropods (this is probably also due to the fact that the animal was walking on all fours, therefore relieving a part of the weight the feet normally supported).
This was a rather small animal (ornithopod?) with a pace of 60 cm and a stride of 105 cm. The four footprints are remarkably similar in length and width, with an average length of 24 cm and an average width of 22 cm (the differences being below 0.5 cm).
The paleoenvironment and its significance
As mentioned before, the paleoenvironment of the area is considered to be deltaic. Reconstructions, based on well logs and sequence stratigraphy, infer a north-east to south-west shoreline (the Western Internal Seaway) with rivers draining towards the south-east.9 Several transgressions (ocean rising and the shoreline moving inland) interrupted the Dunvegan advance to the south-east, resulting in seven marine tongues within the Dunvegan Formation. These episodes are interpreted as tectonically induced subsidence during periods of renewed thrusting in the Cordillera.9 Within a long-age framework, Bhattacharya pointed out that ‘these seven events are beyond the limits of stratigraphic resolution in terms of absolute time and are inferred to have been relatively rapid (a hundred thousand years or so)’.9
The standard ‘long-age’ interpretation of deltaic and near-shore deposits is commonly taught as the process that formed most of the massive sandstone sedimentary sequences around the world. However, such sequences bear little resemblance to the complex and irregular geological profile of modern-day river deltas. The ‘present’ is clearly not ‘the key to the past’ for these formations. A more satisfactory interpretation would be a depositional setting comprising fluctuating floodwaters within a tectonic basin, which was being rapidly filled with sediments washed in from the eroding continent during the Inundatory stage (first half of the Flood).17 Under such circumstances, it is possible that at least some of the dinosaurs recurrently arrived in the area on large floating mats of wood debris from the Flood.

It is interesting to notice that although there was—according to the evolutionary interpretation—no physical barrier to prevent sauropods (the largest of dinosaurs) reaching western Canada during the Cretaceous, no sauropod tracks or bones have been found in Canada thus far.18 The evolutionary explanation for this is ‘ecological reasons’.19 This is a rather fuzzy terminology, since dinosaur remains—including those of large carnosaurs that appear to have fed on sauropods—have been found in nearby polar regions (Alaska). Generally speaking, according to evolutionary interpretations, climate and environmental conditions were favourable for dinosaurs throughout the entire North American continent during the Cretaceous, so the absence of sauropods in the Canadian part of the continent is very difficult to explain in terms of ‘environmental barriers’—especially in the context of abundant theropod fossils. Carnosaurs were quite mobile and they should have followed (like most predators today) the large sauropod herds which supplied a much more easily obtained and more abundant food supply than the more mobile hadrosaurs and the well-protected ceratopsians and ankylosaurs.
Within a young-earth Flood scenario, such an unusual distribution is easy to explain. It is possible that larger dinosaurs like the sauropods did not have the opportunity to find temporary refuge from the floodwaters ‘on board’ floating mats. From a paleoenvironmental point of view, the dinosaur fossil record is mostly located within 200 km of the sea.20 Some isolated specimens were found in marine deposits (Alabama, Kansas, Montana, New Jersey, Alberta, Saskatchewan, England and France) or in hypersaline lagoons (France, Spain and Germany).20 Sometimes, dinosaurs are associated with marine fossils, suggesting nearby shorelines (Mexico, Tanzania and Texas). Even dinosaur fossils found at sites interpreted to be far away from the sea, like the Late Cretaceous ones in Mongolia, are believed to have been buried in sand dunes saturated in rain water.21 All this suggests that the vicinity of the sea, rather than their normal habitat, was where most dinosaurs were buried (with or without transport involved). This is consistent with a global catastrophic Flood scenario. As for the specimens found in marine deposits, while difficult to explain from an evolutionary, global-catastrophe-free scenario, they are perfectly consistent with the biblical Flood!
Fossil-track hunting

Although undoubtedly tempting, the finding of indisputable human footprints synchronous with dinosaur tracks remains an elusive quest. Furthermore, it could be potentially harmful if track finds are not properly dealt with, as has previously happened (see the Paluxy River controversy). I appreciate the way the discoverers of the ichnofossils in the Tumbler Ridge area handled their finds. It was obvious from the pictures they sent to AiG-Canada that they believed they had found human footprints. As enthusiastic Christians and strong believers in the historical narrative of Genesis, they could have broadcast their discoveries to support their position, but they requested a professional investigation before making any announcement. Although my assessment did not confirm their initial suggestion, they not only accepted the verdict against human ‘authorship’ but also actively promoted it and now use their new experience to better educate Christians during their frequent field trips to the sites.
I recommend that everyone actively investigating, or hunting for, human ichnofossils follow several simple, yet necessary, steps in the field:
- Check the parallelism of any track’s bottom plane with the local dip of the host layers. If they are not parallel, discard the item.
- Look for trackways (minimum of two consecutive left-right or right-left footprints). Isolated prints will never suffice to build a case.
- Try to rule out any alternative explanation (like mud collapse, slumping, eroded metatarsal print of a tridactyl animal, etc.).
While in the field, I often recall these words of Sherlock Holmes: ‘Once you eliminate the impossible, whatever remains, no matter how improbable, must be the truth.’ ‘Impossible’ here, however, should not be defined according to a bias (i.e. an evolutionist would claim that it’s impossible for a human footprint to be in the same layer with dinosaur footprints), but it should be the logical kind of ‘impossible’.
Acknowledgments
I wish to deeply thank the Walkleys and the Zimmermans for their friendship, enthusiasm and generosity. Their open-mindedness and dedication were a joy to me, and working with them strengthened my faith and tremendously increased my motivation.
References
- Lockley, M. and Hunt, A.P., Dinosaur tracks and the fossil footprints of the Western United States, Columbia University Press, p. XIII, 1995. Return to text.
- Lockley and Hunt, ref. 1, pp. 11–12. Return to text.
- Lockley and Hunt, ref. 1, p. 38. Return to text.
- Lockley and Hunt, ref. 1, p. 40. Return to text.
- Kvale, E.P. et al., Middle Jurassic (Bajocian and Bathonian) dinosaur megatracksites, Bighorn Basin, Wyoming, USA, Palaios 16:233–254, 2001. Return to text.
- Oard, M., Newly discovered dinosaur megatracksites support Flood model, Journal of Creation 16(3):5–7. Return to text.
- It is likely that many of the species names for dinosaurs are given for the same species with different sizes and shapes, or found in different countries. Return to text.
- Plint, A.G., Sequence stratigraphy and paleogeography of a Cenomanian deltaic complex: the Dunvegan and lower Kaskapau formations in subsurface and outcrop, Alberta and British Columbia, Canada, Bull. Canadian Petroleum Geology 48(1):43–79, March 2000. Return to text.
- Bhattacharya, J.P., Cretaceous Dunvegan Formation of the Western Canada Sedimentary Basin, May 2004. Return to text.
- <www.members.shaw.ca/vertebrateichnology/>, May 2004. Return to text.
- Kuban, G.J., Elongate dinosaur tracks; in: Gilette, D.D. and Lockley, M.G. (Eds.), Dinosaur Tracks and Traces, Cambridge University Press, New York, pp. 57–72, 1991. Return to text.
- Fields, W., Miller, H., Whitmore, J., Davis, D., Detwiler, G., Ditmars, J., Whitelaw, R. and Novaez, G., The Paluxy River footprints revisited; in: Walsh, R.E. and Brooks, C.L. (Eds.), Proc. 2nd Int. Conf. Creationism, Creation Science Fellowship, Pittsburgh, vol. 2, pp. 155–168, 1990. Return to text.
- Kuban, ref. 11, p. 62. Return to text.
- Kuban, ref. 11, pp. 69–71. Return to text.
- Locations include: Connecticut, Colorado, New Mexico, Texas, Australia, South Africa, China, Spain and Morocco (ref. 1). Return to text.
- Lockley and Hunt, ref. 1, p. 126. Return to text.
- Walker, T., A biblical geologic model; in: Walsh, R.E. (Ed.), Proc. 3rd Int. Conf. Creationism, Creation Science Fellowship, Pittsburgh, pp. 581–592, 1994. Return to text.
- Currie, P.J., Dinosaur footprints of western Canada; in: Gilette, D.D. and Lockley. M.G. (Eds.), Dinosaur Tracks and Traces, Cambridge University Press, New York, p. 298, 1989. Return to text.
- Lockley, M. and Conrad, K., The paleoenvironmental context, preservation, and paleoecological significance of dinosaur tracksites in the western USA; in: Gilette, D.D. and Lockley. M.G. (Eds.), Dinosaur Tracks and Traces, Cambridge University Press, New York, pp. 121–134, 1989. Return to text.
- Dodson, P., Dinosaur paleoecology, May 2004. Return to text.
- Achenbach, J., Flash & bone, National Geographic, March 2003, p. 27. Return to text.

Readers’ comments
Comments are automatically closed 14 days after publication.