

Journal of Creation 19(2):109–118, September 2002
Browse our latest digital issue Subscribe
Parkie: a new ‘pseudoplesiosaur’ washed up on the Nova Scotia coast
 Figure 1. Posterior view of carcass.  Figure 2. Side view of head showing nares, eye-socket and empty eyeball (see arrows). 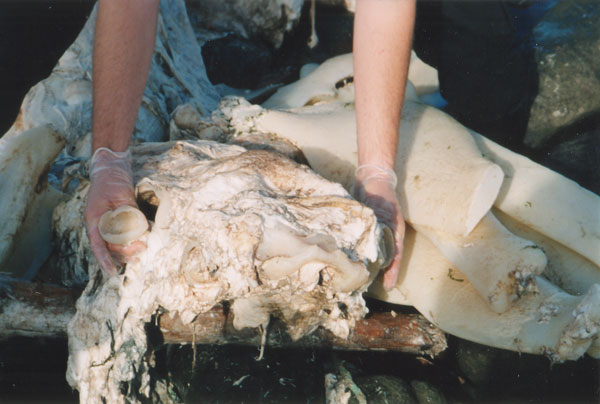 Figure 3. Front view of head next to jaw/gill cartilage.  Figure 4. View of head from above showing two finger-like projections.  Figure 5. Anterior view of carcass showing broken ‘collarbone’ (see arrows).  Figure 6. Tail. |
Abstract
A ‘pseudoplesiosaur’ carcass found on the Nova Scotia coast was thoroughly studied. Its major external and internal features were examined and photographed, and some tissue samples were further analyzed in the laboratory. The carcass was clearly identified as that of a basking shark. This study helped to establish characteristic features of basking shark decomposition that should be useful in identifying these huge creatures that are periodically washed up on coastlines around the world.
Introduction
On 14 September 2002, a large carcass washed up on the shores of Parkers Cove, Nova Scotia, Canada. According to Canadian news:
‘The eight-metre long creature has a small head that attaches to a long thin neck then to a massive body of cavities and cartilage. Huge, empty eye sockets gave the carcass an eerie look. Strangely, long strands of coarse hair cover the fins—a confusing detail.
‘Some say the beast has the makings of a famous sea monster—though the neck may be shorter than what we’re used to with the Loch Ness monster.’1
Many local fishermen had claimed to have seen this creature from time to time, and said it had a ‘horse-like’ head when it peeked out of the water.
The furore surrounding the Parkers Cove beast, Parkie, a phenomenon which always seems to follow the sightings of these ‘monsters’, resulted in Canadian universities and scientists being accused of not showing any interest in this potentially earth-shattering find—most universities simply dismissed this find as a basking shark without any further analysis. However, Professor Herman and his colleague Dr Don Stewart from Acadia University, Nova Scotia, volunteered to carry out DNA analysis on a tissue sample from Parkie.
A preliminary report on this carcass was posted on the web,2 but it did not properly address all the issues and contained many errors. We therefore decided to do a thorough study of Parkie, especially since this carcass showed an uncanny resemblance to the creature known as the Zuiyo-maru carcass (ZMC) which was hauled up by a Japanese fishing boat (the Zuiyo-maru) off the New Zealand coastline in 1977,3 and to the Kaikoura-1 and Kaikoura-2 carcasses washed up on the Kaikoura coastline of New Zealand.4,5
One of us visited the site of the carcass on 18 September 2002, and was initially impressed by the plesiosaur likeness of Parkie. Its major external and internal features were recorded and photographed, and some tissues were later analyzed in the laboratory. A careful study of all the evidence has now helped to shed more light on the nature of these creatures found washed up around the world.
Parkie and ZMC similarities
On first observation, Parkie appeared solid and mostly intact, although much of the skin, and what initially appeared to be its throat and part of the tail were missing. It also demonstrated an uncanny similarity to the 1977 ZMC. A comparison of some structures/features of ZMC and the measurements made by Michiko Yano closely matched those of Parkie (see figure 1 and table 1).
Also initially noticeable was a very strong cod-liver-oil-like smell. This was also mixed with a strong putrid/nauseating smell typical of dead animals. There was, however, no ammonia smell such as would be expected from a rotting fish.
The skeleton of the carcass was typical of washed up ‘pseudoplesiosaurs’ and was made entirely of cartilage. The head/skull was quite hard and featured the typical ‘nare’-like structure at the front (figs 2 and 3). It was also rounded, not unlike the ‘head of a turtle’, and remarkably similar to the photograph and the drawing by Yano of ZMC.5 Further observation of the skull revealed what appeared to be two eye sockets with the remains of eyeballs hanging out of them and attached to optic nerves (figs 2 and 3). There were also two 150 mm finger-like cartilaginous projections, one above each ‘nare’ (fig. 4).
The body proportions and shape were very similar to those of ZMC (see table 1 and figure 1). Parkie was 8 m long, with a 1.37-m-long neck (fig. 5), and a 0.7-m-long tail (fig. 6)—the Kaikoura-2 carcass measured a comparable 8.8 m in length.
There was a large collarbone/pectoral girdle which was broken near the neck (fig. 5 and 7). This structure appears identical to that of ZMC and, as already mentioned, is the wrong shape for a plesiosaur which has a flat body plan.5 The collarbone could have broken during contact with the rocks on the beach at Parkers Bay. In comparison, ZMC was fished up from the sea bottom off the Banks Peninsula, Christchurch, and therefore its skeleton appears to have been more protected against damage.
Table 1. Overall measurements of ‘sharkosaur’ carcasses. ND = Not determined. |
|||
| Measurements of organs | ZM carcass3 | Parkie | Kaikoura-24 |
| Overall length | 10 m | 8 m | 8.8 m |
| Body length | 6 m | 5.9 m | ND |
| Skull length | 450 mm | approx. 450 mm | ND |
| Skull width or ‘front view’ | 300 mm | approx. 350 mm | ND |
| Neck length | 1.5 m | 1.37 m | ND |
| Tail length (+ number of vertebrae) | 2 m (15 vertebrae) | 0.7 m (7 vertebrae) | (13 vertebrae) |
| Neck vertebrae diameter | 200 mm | approx. 180 mm | ND |
| Back vertebrae diameter | 150 mm | ND | ND |
| Tail vertebrae diameter | 125–130 mm (at base of tail) | 140 mm | ND |
| ‘Rib’ length | 400 mm | ND | approx. 400 mm |
| Pectoral fin length | 0.98 m | 1.07 m | ND |
| 1st Dorsal fin length | ND | approx. 610 mm | Not present |
| Pelvic fin length | ND | 305 mm | ND |
| Clasper length | ND | 0.58 m | ND |
| ‘Horny fibre’ length (pectoral fin) | 200–300 mm | 152–203 mm | ND |
| ‘Horny fibre’ diameter | 2.5 mm | 2.5 mm | ND |
The tail and neck vertebrae of Parkie (figs 6, 8 and 9) were almost identical in shape and size to that previously reported for the ZM and the Kaikoura-1 and -2 finds (see fig. 10): they were block-shaped and without vertebral processes, which is typical of sharks and other fish but not of a tetrapod such as a plesiosaur.
 Figure 7. Broken cartilage ‘collarbone’.  Figure 8. Neck vertebrae.  Figure 9. Close up of neck vertebrae showing horse-like mane (see arrow). Photo by Bev Elliot  Figure 10. Posterior view of Kaikoura-1 find.  Figure 11. Pectoral fins.  Figure 12. Posterior view showing connective tissue. |
The tail was shorter than that of ZMC—7 vertebrae compared to 15 (figs 1 and 6)—but it did not taper off at the end like that of ZMC, suggesting that a segment with smaller vertebrae had broken off. Tails breaking off and the loss of the tail fin are well-known characteristics of basking shark decomposition.6
The one-metre-long pectoral fins were also attached to the middle of the pectoral girdle (figs 5 and 11). They had the same shape and proportion to the rest of the body as the fins of ZMC. Although these fins were wide/broad at their base, they were attached at a narrower point like those described by Yano for ZMC (see fig. 3 in ref. 4). It is important to note that Yano’s drawing of the pectoral fins looks nothing like his photographs.4
There was also a pair of rear, or pelvic, fins which were clearly smaller than the pectoral fins (figs 1, 12 and 13). Although Yano drew both pairs of fins the same size, he did not take a picture of the rear fins, so his claim cannot be supported. It is important to note that in an interview, Yano mentioned that the rear fins could have been smaller: ‘“How about the size of the front and back fins,” Obata asked. “I don’t think there was much difference,” Yano said. “If I try to remember, I think the front was bigger … I regret you can’t see this well from the picture”’7 [emphasis added].
All of Parkie’s fins featured the characteristic pseudoplesiosaur’s horny fibres or ceratotrichia around the fin edges (figs 11 and 13).
About 50% of skin was missing and most of the surface of the carcass was whitish (figs 1, 5 and 11) like ZMC. Some remaining grey skin was present on areas such as the pectoral fins (see below). Some black tufts of hair-like fibres were also visible, especially around the neck area where the carcass was partly covered with what appeared to be a motley mane (figs 8 and 9). This is also on par with ZMC and other washed up pseudoplesiosaurs, and is due to partial fraying of surface muscle.
The carcass had mainly white muscle with a chicken breast consistency. According to Yano, ZMC muscle was also mainly white except that ‘Reddish muscles were observed around the caudal vertebrae when the tail was partly cut near its base.’8 Parkie likewise featured some red muscle, but this was limited to the spinal area, which also includes the tail.
Features identifying Parkie: tetrapod or fish?
Parkie’s white muscle had strong bands of elastic connective tissue, which appeared identical to the mycommata anchoring the muscles of fish and sharks (figs 1 and 12). This was also observed for ZMC, but is not a characteristic of tetrapods.
Underneath the ventral muscles, a 150–200 mm layer of fat tissue protected the internal organs (figs 14 and 15). There was, however, no rib-cage as would be expected for tetrapod—both ZMC and the Kaikoura-2 carcass also had no proper ribs—which again is consistent with a fish/shark identity.
Underneath Parkie’s long tail, there was a highly decomposed anal fin (fig. 16). Such a fin is clearly unlike that of any known tetrapod. A more careful observation of all the fins revealed that they did not contain any bone, but were mainly made up of connective tissue, dermal fibres and cartilage (figs 13 and 16). This is also a typical characteristic of sharks and other fish.
Immediately behind the rear fins there were two long, hard, cartilaginous appendages with claw-like endings. These were in the same anatomical position and had the same shape and proportions as expected for sexual claspers—mating structures which are only present in the pelvic fins of male sharks (figs 1, 12, 13 and 17).9 From their appearance, it is obvious how these structures could have easily been confused with an extra set of flippers/fins or appendages in washed-up ‘monsters/serpents’.
A structure at the centre of much debate, a dorsal fin, was also clearly present. A single fin was attached to the mid-dorsal section of Parkie, a position expected for a fish or shark (see figs 5 and 18). This fin was noticeably smaller than the pectoral fins (fig. 11), but more triangular. It also featured a free rear tip near its base, a characteristic of sharks. Like the other fins it had horny fibres, but mainly on the posterior edge facing the rear of Parkie up to the fin apex. Cartilage and connective tissue connected it to the torso.
 Figure 13. Close-up of pelvic fin and partial view of clasper.  Figure 14. Fat tissue covering internal organs.  Figure 15. Internal organs including two lobes of the liver.  Figure 16. Anal fin.  Figure 17. Clasper (see arrow).  Figure 18. Dorsal fin showing free rear tip. |
Looking closer at the much-debated picture of the back of the ZMC (fig. 19), it is evident that there is an identical dorsal fin at the same position. The first dorsal fins of Parkie and ZMC are clearly similar in size as well as in shape and overall proportions. They both also have strand-like connective tissue at their base and horny fibres along the posterior edge and apex of the fin. The ZMC fin appears more rounded at its apex, but this may be due to a difference in its level of decomposition (and possibly variation between basking sharks populations in New Zealand and northern US/Canadan).10 It is also gradually detaching from the torso, a characteristic of basking shark decomposition.4 Because of its small size compared to the rest of the carcass and the more prominent pectoral fins, Yano appears to have missed this in his description of the ZMC. What probably also helped him overlook this fin is that it had considerably deteriorated and did not have the typical shape of a shark fin.
Careful observation of the underside of parkie’s skull revealed palate rills, indicating that this was the upper palate of the creature (fig. 2) and that the rest of the mouth and bottom jaw had detached from the head—another well-known feature of the decomposition process of basking sharks.11 The case for the missing mouth parts was strengthened when a number of long cartilagenous structures were identified 6 m away from the carcass—they were narrow, long and curved, and appeared to be part of the displaced jaw and gill structures of the creature (figs 3, 20 and 21).12
A 15–20-cm layer of fat was removed to uncover the internal organs. The organs appeared to be all intact in contrast to those described for the ZMC, which were damaged and eaten by worms/fish.13 Since the internal organs of these creatures had not been described in the previous carcasses, we were fortunate to be able to study them here.
A number of organs/tissues could be identified, including a very long stomach. Among the most prominent structures were two very long organs, measuring 3.66 m and 3.2 m, respectively (figs 14 and 15). These are consistent with the livers of sharks, which are made up of a small central or median lobe, and two large lobes which can be up to a third of the shark’s body length,14—if we add the length of the missing tail and head parts to Parkie’s 8 m carcass, the liver would be about one third of its length. In basking sharks the liver is typically 20% of its body weight,15 and these sharks are today still hunted commercially because of the large stores of oil in their livers.16 This also helped to explain why the carcass had such a strong cod liver oil smell.
Skin analysis
The skin colour of Parkie, from areas where the skin was still predominantly intact, was greyish-light brown and had a characteristic sandpaper feel of shark skin (fig. 11). This colour is a match for basking sharks, which usually have a greyish brown to slate grey, or almost black, upper surface.15
A sample of skin tissue was removed and later analyzed by electron microscopy. The fine structure of the skin was typically shark-like and made up of small barbs, known as placoid scales or dermal denticles, which are responsible for the sandpaper characteristic of shark skin (figs 22 and 23).17 A hollow interior could also be seen in some of the broken denticles. This is consistent with the pulp cavity of denticles (fig. 24) and further confirms that these structures were indeed vascular (supplied with blood) shark denticles. Parkie’s barbs, however, were conical with a pointy apex and quite distinct from the flatter denticles of other sharks such as the whale shark (the world’s largest), white shark, etc. (fig. 25). Also, unlike these other sharks, the denticles pointed in at least three directions (see figs 22 and 23). Denticles that point in all directions instead of uniformly tailward is a distinct feature of basking shark skin.17
DNA sequencing
Courtesy Todd Wood Figure 19. View of Zuiyo-maru carcass showing dorsal fin.
Professor Herman and Dr Don Stewart from Acadia University obtained a tissue sample from Parkie, from which they extracted some DNA and carried out PCR (Polymerase Chain Reaction) analysis using some basking-shark-specific DNA primers, ‘bscythF2’ and ‘bscytbR1’.18
| Primer | Primer DNA Sequence |
| bscytbF2 | 5’ CGTAGGCTATCTTTTGCC 3’ |
| bscytbR1 | 5’ GTGATTAGGAAGGGGAGA 3’ |
The primers had been developed by a UK laboratory to help check for basking shark products and derivatives in commercial products, because of concerns of a worldwide decline in basking shark numbers. These primers have been shown to have a high specificity for basking shark DNA as they need to stand up to legal scrutiny. The primers are based on Cetorhinus maximus (basking shark) cytochrome b (cytb) gene, a mitochondrial gene which encodes mitochondrial protein.
 Figure 20. Jaw/gill structures. 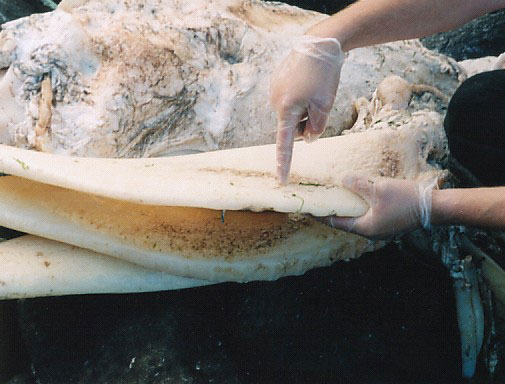 Figure 21. Close up of a jaw/gill structure. Photo by Mark Armitage  Figure 22. Highly magnified denticles on Parkie’s skin (SEM micrograph, courtesy Mark H. Armitage M.Sc., ICR EM Lab). Photo by Mark Armitage  Figure 23. Broken denticles showing pulp cavity (SEM micrograph, courtesy Mark H. Armitage M.Sc., ICR EM Lab). |
According to the strategy, if the PCR results proved to be negative, this would mean that Parkie was not a basking shark, and further analysis would need to be carried out to try to identify what type of creature it was. According to Herman:
‘When tested, the samples were consistently and unequivocally positive. The DNA amplified very strongly, indicating a match with Basking Shark. There is now little doubt in my mind, based on the DNA evidence, that Parkie was indeed a Basking Shark.’19
In order to double check that the amplified DNA fragment did indeed correspond to cytb gene, its DNA sequence was analyzed. The sequence/procedure was able to resolve 148 out of the 186 bases (80%) of the amplified fragment (fig. 26).20,21 A BLAST search (Basic Local Alignment Search Tool; a method for rapid searching of nucleotide and protein databases) with this sequence most closely matched that of basking shark cytb gene, with 146 matching bases out of 148, i.e. 99% similarity.22
It was also interesting to note that the seven most similar sequences in the BLAST search were cytb genes from other sharks—i.e. sequences from 123 down to 119 bases in length, with 89% to 87% identity—such as Longfin mako, great white shark and big-eye thresher shark. The PCR analysis therefore unequivocally confirms a basking shark identity for Parkie.
The amplified sequence differed from the published cytb gene sequence only in two positions, each with a ‘T’ (thymine) instead of a ‘C’ (cytosine). The change at nucleotide position 458 corresponds to a silent/neutral change, as both codons (nucleotide triplets coding for an amino acid), ggc and ggt, code for the amino acid glycine. The second change at nucleotide position 493, from ctc to ttc, however, corresponds to a change from the basic amino acid histidine to the non-polar phenylalanine.
These two nucleotide mismatches correspond to a 1.4% difference in sequence, which is comparable to the 1.1% difference observed among basking sharks in the 188bp region further downstream in the cytb gene (see highlighted sequence from nucleotide numbers 707 to 908 in figure 27), and may likewise represent variation between the two known basking shark haplotypes/variants of the cytb gene.23
Discussion
The results of this study clearly show that Parkie is the same kind of creature as the one hauled up by the Zuiyo-maru trawler off the New Zealand coast in 1977, (and for that matter the Kaikoura carcasses too). The two carcasses show an uncanny similarity, sharing the same body shape, proportions and general size. They both sport a cartilaginous skeleton lacking a rib cage, block-shaped vertebrae without vertebral processes, fish-like fins (not flippers), as well as white muscles with strong elastic connective tissue. These are all characteristic of fish but not tetrapods such as mammals or plesiosaurs. Moreover, they both share identical features expected for a basking shark such as size (second-largest fish after the whale shark), jaws/gills (mouthparts) that fall off during decay, a typical shark dorsal fin (with a free rear tip on Parkie), and the presence of horny fibres on the fins. What unequivocally characterizes Parkie as a basking shark—and thus also ZMC and both Kaikoura finds—are its two huge liver lobes, a pair of claspers (making Parkie a male shark), skin denticles that point in all directions, and finally basking shark specific mitochondrial DNA.
As previously documented, all the decomposing basking shark carcasses display the same tell-tale characteristics, such as: the loss of gill-arches and jaw parts, leaving a turtle-like cranium; the loss of the caudal fin; a ‘mane’ resulting from fraying muscle; fins with horny filaments; and mycommata (connective tissue anchoring the muscles of fish and sharks). From our study of Parkie and ZMC (and the Kaikoura-2 carcass) we have now also identified additional features of these decomposing pseudoplesiosaurs, such as: nare-like structures which are simply part of the cranium; a pair of finger-like projections from the cranium; what appear to be large openings/orbits for the eyes; a visible large pelvic girdle; and the early loss of the dorsal fin.
The overall conclusion of the original CPC research report, together with the findings of our previous study,5 clearly supported a basking shark identity for the ZMC. One of the CPC studies pointed out a number of apparent basking-shark inconsistencies5,24 and some have claimed that these constitute sufficient evidence for ZMC being some type of unknown mammal or plesiosaur-like creature.25 However, these features have now been found in Parkie:
(a) ‘The covering of strong dermal fibres—as in mammals.’ These were clearly present on Parkie as grey/black tufts and ‘mane’, and are a result of fraying of muscle tissue.
(b) ‘The fat-like tissues—fat is not found in fish.’ We found a 150–200 mm layer of fat tissue over the white muscle around the ventral area, which covers the internal organs. Moreover, basking sharks are known for having large fat deposits in their skin.13
(c) ‘The red muscles—not possessed by fish.’ Red muscle was clearly evident along the spinal area of Parkie.
 Figure 24. Cross-section of a shark dermal denticle (after Springer and Gold).28  Figure 25. Shape and arrangement of typical shark denticles. |
(d) ‘The smell was of a mammal, not the strong ammonia smell of putrefying fish and sharks.’ Parkie did not smell of ammonia either, but rather had a strong putrid/nauseating smell, like that of a dead land animal, mixed with a strong cod liver oil smell.
(e) ‘The head was hard, unlike that of a fish.’ This also matches Parkie’s cranium.
(f) ‘The nares were on the front of the skull—not like sharks.’ These structures were present in Parkie.
All these characteristics of ZMC and Parkie can therefore simply be added to the list of typical features for basking shark carcasses.
Another feature also regularly brought up against the basking shark identity of ZMC is the size and composition of the pectoral fins. Yano drew both the pectoral and pelvic fins the same size. But he did not take a picture of the rear fins, and during an interview, he mentioned that the rear fins could have been smaller. So this cannot be considered as evidence. Although Yano also felt that the pectoral fins consisted of bone, this observation carries no weight, as the hard cartilage in basking shark fins would give the same impression when ‘trod on’ (see A ‘tail’ of many monsters on p.74–75).

Figure 26. Partial coding sequence of Cetorhinus maximus (basking shark) cytb gene.29 Sequences for primer bscytbF2 (5’ to 3’) and complimentary sequence for primer bscytbR1 (3’ to 5’) are in bold and underlined. Sequence determined from amplified DNA fragment in figure 26 (from nucleotides 342 to 527) is shown in bold. The two mismatches (‘c’ instead of ‘t’) at nucleotides 459 and 493 are capitalized and underlined, and the resulting new 3-base codons are also underlined. A second 202 bp sequence further downstream is marked in bold.
Much has also been commented about ZMC’s lack of a dorsal fin, or even the belief by some of a symmetrical pair of small dorsal fins.25,26 But in Yano’s photograph, ZMC clearly has a comparable mid-dorsal fin to Parkie’s (fig. 19). The ZMC fin appears more rounded at its apex, but this may simply be a result of further decomposition and possibly also variation between basking sharks populations in New Zealand and northern US/Canada. The differences noticed in the measurements of the fins, vertebrae, etc., may also be due to differences between species or simply due to sex variations of basking sharks.
Yano did not mention or draw any claspers on ZMC either. Since these prominent structures are closely associated with the pelvic fins of male sharks, the ZMC appears to have been that from a female basking shark.
From this study, Parkie appears to fit the description of many of these washed-up creatures, indicating that they are simply basking sharks. It is also interesting to note that these ‘pseudoplesiosaurs’ frequently wash up in areas where basking sharks live, the world’s arctic and temperate waters which include the coasts of eastern Canada and New Zealand.12
All the locals of Parkers Cove rejected outright the idea of Parkie being a basking shark. But this is nothing new, it appears many have been fooled by the similarity of basking shark remains to a plesiosaur.27 If there is one thing we can learn from the study of these carcasses, it is that we should be careful to draw conclusions from only visual observations of the remains of decomposing creatures.
References and notes
- CTV News Staff, Washed up creature on N.S. beach stumps locals, 17 September 2002; www.ctv.ca/servlet/ArticleNews/story/CTVNews/20020917/fundy_shark020917/SciTech/story/. Return to text.
- Sundberg, J. and Buccola, A., The Nova Scotia final report versus an Anthony Buccola summary, 2002; www.bahnhof.se/~wizard/cryptoworld/index215a.html, 8 September 2004. Return to text.
- Collected papers on the carcass of an unidentified animal trawled off New Zealand by the Zuyo-maru, Sasaki, T. (Ed.), La Société Franco-Japonaise d’Océanographie, Tokyo, pp. 45–83, 1978. Referred to as ‘CPC’. Return to text.
- Jerlström, P. and Elliott, B., Letting rotting sharks lie: further evidence for shark identity of Zuiyo-maru carcass, TJ 13(2):83–87, 1999. Return to text.
- Jerlström, P.G., Live plesiosaurs: weighing the evidence, TJ 12(3):339–346, 1998. Return to text.
- www.fao.org/figis/servlet/FiRefServlet?ds=species&fid=2005, 14 September 2004. Return to text.
- Koster, J., Creature feature, Oceans 10:56–59, 1977. See www.gennet.org/nessy.htm, p. 2. Return to text.
- Obata, I. and Tomoda, Y., Comparison of the unidentified animal with fossil animals; in: CPC, ref. 3, p. 49. Return to text.
- www.amonline.net.au/fishes/fishfacts/fish/cleucas.htm, 14 October 2004. Return to text.
- www.elasmo-research.org/education/shark_profiles/cetorhinidae.htm, 14 October 2004. Return to text.
- www.gma.org/fogm/Cetorhinus_maximus.htm, 14 September 2004. Return to text.
- www.flmnh.ufl.edu/fish/Gallery/Descript/baskingshark/baskingshark.html, 16 September 2004. Return to text.
- Kuban, G.J., Sea-monster or Shark? An analysis of a supposed plesiosaur carcass netted in 1977, Reports of the National Centre for Science Education 17(3):16–28, 1997; paleo.cc/paluxy/plesios.htm, 15 September 2004 . Return to text.
- www.pc.maricopa.edu/Biology/ppepe/BIO145/bio_images/skvisce2.gif, 16 September 2004. Return to text.
- www.elasmo-research.org/education/shark_profiles/cetorhinus.htm, 16 September 2004. Return to text.
- www.fao.org/figis/servlet/FiRefServlet?ds=species&fid=2005, 16 September 2004. Return to text.
- www.amonline.net.au/fishes/students/scales/placoid.htm16 September 2004; www.elasmo-research.org/education/white_shark/scales.htm, 16 September 2004. Return to text.
- Hoelzel, A.R., DNA identification of basking shark Cetorhinus maximus products in trade: Report submitted by the United Kingdom in support of proposal 11.49 to add Cetorhinus maximus to Appendix II of CITES, 21 May 2003. Return to text.
- Tom Herman personal communication, 23 May 2003. Return to text.
- Note that it is often difficult to sequence nucleotides proximal to the sequencing primer, so in this case the primer bscytbF2 was most probably used to do the sequencing. Return to text.
- Don Stewart personal communication, 11 July 2003. Return to text.
- www.ncbi.nlm.nih.gov/blast/Blast.cgi, 27 September 2003, RID=1061958952–5830–21796.BLASTQ3. Return to text.
- Hoelzel, A.R., Shark fishing in fin soup, Conservation Genetics 2:69–72, 2001. Return to text.
- Obata, I. and Tomoda, Y., Comparison of the unidentified animal with fossil animals; in CPC, ref. 3, p. 48. Return to text.
- Bowden, M., The Japanese carcass: a Plesiosaur-type Mammal! A Review of the Evidence, Creation Science Movement, Portsmouth, England, p. 4, 2001. Return to text.
- Goertzen, J., New Zuiyo maru cryptid observations: strong indications it was a marine tetrapod, CRSQ 38(1):19–29, 2001. Return to text.
- See refs. 5 and 14. Return to text.
- Springer, V.G. and Gold, J.P., Sharks in Question: The Smithsonian Answer Book, Smithsonian Institution Press, Washington, DC., 1989. Return to text.
- From SRS @EMBL-EBI, www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=Nucleotide&list_uids=07259785&dopt=GenBank, 22 May 2003. Return to text.
Henry de Roos received his Bachelor of Science (1987) in Crop Science and his Masters degree in Agriculture Science (1989) at the University of Guelph, Canada. He also received his Bachelor of Education (1990) from the University of Western Ontario. He worked in biochemical research for a chemical company for 1 year and taught high school science for 6 years. Adept at doing creation evangelism in public schools, he has served with Open Air Campaigners Canada for the last 9 years (he is currently National Director). Henry was one of the first speakers for what is now CMI Canada. Return to text.
Readers’ comments
Comments are automatically closed 14 days after publication.