

Journal of Creation 20(1):7–8, April 2006
Browse our latest digital issue Subscribe
The origin of bubonic plague
Although some forms of the bacterium Yersinia are harmless, other forms have devastated human populations, causing a plague of biblical proportions (Psalm 91:3-7, Psalm 91:9-10,). Bubonic plague, also known as the ‘Black Death’ that killed one fourth of Europe’s population in the 1300s, appeared as a great pestilence several times in the Old Testament, including in Psalm 91 and in 2 Samuel 24:14-25. Perhaps the clearest example of such a plague is recorded in 1 Samuel 6:4-19, where there is a specific reference to the tumors on people (bubos = the tumors of lymph glands) and to rats (the animal vector that carried the plague bacterium, Yersinia pestis.) The biblical time frame for the plagues described in 1 Samuel was about 3,000 years ago.1
Interestingly, experts on plague ‘evolution’ estimate the emergence of Y. pestis at about 1,500-20,000 years ago (within an evolutionary timeframe, of course).2
Plague’s origin is multifaceted
Many infectious diseases can be traced back to the decay and corruption of the original created design of microorganisms as a result of the Fall. Corruption literally means to destroy (from the Latin corruptus). The origin of pathogenic (disease-causing) bacteria such as Y. pestis is complex and multifaceted, and may be explained by a combination of genes that were lost, added and moved. The story of Yersinia’s degeneration into the plague pathogen may serve as a model of ‘fast’ genomic decay and corruption.2
It appears that the beginning of pathogenicity in the genus Yersinia started with a net loss of chromosomal DNA from its original ‘kind’ (figure 1). Later, there were minor additions of plasmid DNA3 as well as DNA from viruses and other bacteria. A few plasmid genes for toxins (table 1) have been acquired from another existing species, but many chromosomal genes have been lost. It takes only a few such genetic changes to produce a new, extremely infectious variant,4 so it may have taken only hundreds or a few thousand years to produce the current bubonic plague strain that has existed for about the last 500 years.
Loss of chromosomal DNA

Researchers hypothesize that key chromosomal genes (i.e. involved in metabolic pathways) were inactivated/lost in changing from a soil-inhabiting Yersinia to a pathogenic Yersinia species.2 Pathogenic Yersiniae have lost structural information and function in about 149 genes. Of these, 58 are the result of frameshift mutations,5 32 have undergone deletions, and the rest are nonsense mutations.6 These incomplete/inactivated genes or ‘pseudogenes’ are an important feature of the Y. pestis genome.7 Wren2 suggests that the genes lost in Y. pestis affected bioenergetic functions, including dicarboxylic amino-acid metabolism. This reduction of metabolic pathways may have allowed the bacterium to conserve energy. The newly emerged strains (variants) were thus streamlined, which might have contributed to the development of pathogenicity (i.e. plague) due to the genes they lacked. The absence of important biosynthetic genes is believed to be a hallmark of genome decay.
Genes added and moved
The corruption by three genes of a relatively benign recent ancestor of Y. pestis may have played a key role in the emergence of bubonic plague. Hinnebusch and colleagues, a plague expert team at the National Institutes for Health,8 maintains that the acquisition of two plasmid genes (i.e. just a few discrete genetic changes) in recent times changed the fairly harmless, Y. pseudotuberculosis, that causes mild food poisoning, to the agent of the ‘Black Death’. A third gene (carried on plasmid pMT1) produces murine toxin, an enzyme required for the initial survival of Y. pestis bacilli in the flea midgut (table 1).7 By acquiring this last gene from another organism, Y. pestis made a crucial shift in its host range, allowing it to survive in fleas, and devolved to relying on its blood-feeding host for transmission. This is just another example of the flexibility of many microbes in sometimes repackaging themselves into more dangerous agents of infectious disease.
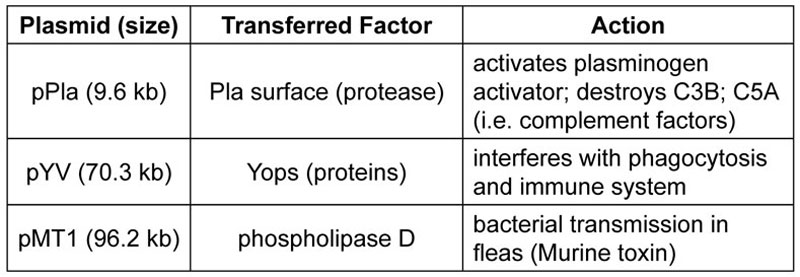
This last corruption is one that distinguishes Yersinia pestis from all closely related, more benign bacteria such as Y. pseudotuberculosis and other Yersinia (e.g. Y. entercolitica).7 In turn, as Y. pestis adapted to rely on its new blood-feeding host for transmission, the emergence of more deadly bacterial strains would have been favoured. It appears that these minor plasmid additions10 were the last changes made in an otherwise long series of genetic losses in Y. pseudotuberculosis’ chromosome (figure 1).
One pathogenicity island was acquired by Yersinia pestis from a different bacterium.2 This cassette of genes was not the result of evolution of new chromosomal DNA, but was an acquisition through lateral gene transfer.11 It produced a corrupted message that gave bacteria a new ‘position’ in the gut. Y. pseudotuberculosis, which lacks the hms locus gene inhabits harmlessly the midgut of the flea. Plague bacilli, by contrast, have this inserted locus gene. Free from their original control, causing a lack of ‘good’ ‘direction’ information, the bacteria migrate from the midgut to the foregut, forming a plug of packed bacilli which is passed on to the victim when the flea feeds.
Genes, germs and Genesis
Plague bacteria are not the only microorganisms that have degenerated into disease-causing organisms. A more common recent example of a harmless bacteria ‘devolving’ into a pathogenic one is the intestinal Escherichia coli O157H7 strain that occasionally causes fatalities.12 Other pathogenic bacteria that have undergone genomic decay include various mycoplasmas (e.g. Mycoplasma genitalium and M. pneumonia, the later causing pneumonia), and Mycobacterium leprae (the leprosy bacillus).13,14
As we study the origin of infectious disease from a creationist, biblical perspective, bacteria provide us with a model of what may have happened to living things over time in a fallen, cursed and corrupted world. Many illnesses can probably be traced back to a loss of genetic information, plasmid acquisition, and gene translocation in organisms such as bacteria, fungi, etc. For those who know the Creator, we can rejoice that someday the Great Physician will restore all plagued bodies to a very good condition once again (Revelation 22:2-3).
Re-posted on homepage: 19 February 2016
References and notes
- Ryrie, C., Ryrie Study Bible (KJV), Moody Press, Chicago, IL, 1994. Return to text.
- Wren, B.W., The Yersiniae-a model genus to study the rapid evolution of bacterial pathogens, Nature Micro. Rev. 1(1):55-64, 2003 | PMID:15040180. Return to text.
- Plasmids are circular, double-stranded units of DNA that replicate within cells independently of the chromosomal DNA. Return to text.
- Lorange, E.A., Race, B.L., Sebbane, F. and Hinnebusch, B.J., Poor vector competence of fleas and the evolution of hypervirulence in Yersinia pestis, J. Infect. Dis. 191(11):1907-1912, 2005 | PMID: 15871125. Return to text.
- Frameshift mutations are changes in DNA where insertions or deletions of sequence occur that are not a multiple of three base pairs, thus disrupting the gene/protein normal code. Return to text.
- A nonsense mutation is any alteration of DNA that causes a codon representing an amino acid to be replaced by a termination codon. Return to text.
- Hinnebusch, B.J., Rudolph, A.E., Cherepanov, P., Dixon, J.E., Schwan, T.G., and Forsberg, A., Role of Yersinia murine toxin in survival of Yersinia pestis in the midgut of the flea vector, Science 296(5568):733-735, 2002 | PMID: 11976454. Return to text.
- Nester, E.W., et al., Microbiology: A Human Perspective, 4th ed, WCB McGraw-Hill Publishers, Boston, MA, 2004. Return to text.
- Parkhill, J., et al., Genome sequence of Yersinia pestis, the causative agent of plague, Nature 413(6855):523-527, 2001 | PMID: 11586360. Return to text.
- Hinnebusch B.J., Perry R.D. and Schwan T.G., Role of the Yersinia pestis hemin storage (hms) locus in the transmission of plague by fleas, Science 273(5273):367-370, 1996 | PMID: 8662526. Return to text.
- Lateral gene transfer is any process in which an organism transfers DNA to another cell that is not its offspring. Return to text.
- Pommerville, J.C., Alcamo’s Fundamentals of Microbiology, Jones and Bartlett Publishers, Sudbury, MA, 2004. Return to text.
- Wood, T.C., Genome decay in the Mycoplasmas, Acts and Facts Impact 340:1-4, 1981. Return to text.
- Eiglmeier, K., The decaying genome of Mycobacterium leprae, Lepr. Rev. 72(4):387-398, 2001 | PMID:1826475. Return to text.



{kind=link}
Readers’ comments
Comments are automatically closed 14 days after publication.