

Journal of Creation 14(3):118–127, December 2000
Browse our latest digital issue Subscribe
Neandertal speech capability and the limitations of osteological analysis
Attempts to show whether Neandertals were capable of articulate speech have been contested on methodological grounds. Much of the debate arises from the question of how much soft tissue information one can glean from Neandertal skeletal remains. This study compares the features of three archaic human vocal tracts with those of modern Homo sapiens by analysing corresponding cervical vertebrae and basicranial heights. The results predict that the La Chapelle Neandertal had a vocal tract like that of modern humans, while the Shanidar-1 Neandertal and the archaic modern human from Skhúl did not. The results suggest a high degree of vocal tract variability in the genus Homo. An observation is made regarding the limitations of anthropometric studies of archaic populations.
History
Much discussion surrounds a series of articles published by Lieberman and Crelin,1 Laitman et al.,2 Lieberman,3 Crelin4 and Lieberman et al.5 on the Neandertal vocal tract and its anatomical implications for articulate speech. The above claim that Neandertals were incapable of producing some modern human vowel sounds, namely English ‘high vowels’ as found in cake, root and eat. Lieberman and Crelin first argue this in terms of the Neandertal basicranium*, among other anatomical features, and its relation to speech anatomy. Neandertals are suspected of having a flatter cranial* base than modern humans. They argue that cranial ‘flexion’, as it is sometimes known, is highly correlated with a high larynx* position in the throat. This trait is observed in newborn humans and chimpanzees, two groups who cannot produce high vowel sounds. Lieberman and Crelin’s peculiar application of recapitulation theory* postulates that Neandertal speech ability was abridged over that of adult modern humans, whose basicrania* are more angled and whose larynges are supposedly lower in the throat. This basicranial flattening correlation with high larynx position is further developed in subsequent papers.2,6 Phylogenetic variability in cranial base flatness has been observed, and based upon Lieberman and Crelin’s model, significant differences in vocal tract height are proposed through attempts at Neandertal soft tissue reconstruction.
* Terms marked with an asterisk are defined in the Glossary at the end of this article.
The cranial base

The basicranial flattening argument is the most thoroughly contested idea regarding Neandertal speech capability. At the heart of the controversy is whether or not the basicranium is a good indicator of larynx and hyoid* bone position. It is argued that Laitman’s method of measuring the cranial base is an unconventional way of establishing flexion angle,7 since Laitman incorporates palatal dimensions and other splanchnocranial* landmarks as well. But others8-11 have not shunned to use this method, even in refuting the assertions of Lieberman, Crelin and Laitman et al. Others have contested that basicranial flattening is common in adult modern humans too, with many displaying flattening on the scale of Neandertals, yet showing no substantial difference in larynx height.10-13 In fact, an extension of the Lieberman, Crelin and Laitman et al. method to modern-day and Mesolithic specimens argues that the high, ape-like larynx position for the La Chapelle-Aux-Saints Neandertal is unsupportable.9 Additionally, the recent La Chapelle cranium reconstruction14 revises the cranial base angle to 137.5°, a figure well within the range of modern human variation. Heim concludes that ‘our reconstruction then should put an end to the controversies about the existence of articulated language among Neandertals. These humans were anatomically capable of producing the same vowels and the same consonants as us’.15 Heim’s 1989 reconstruction of the La Chapelle Neandertal cranium, as well as his very able interpretation of the La Ferrasie Neandertal reconstruction16 as being more modern in appearance, should have put much of this to rest. But it hasn’t. Certainly, the ability to discern larynx position from cranial base studies alone is highly questionable.
Lieberman17 still argues that basicranial flexion is an important component in laryngeal anatomical positioning. Independent studies do show that artificially altering basicranial angle affects ontogenetic* migration of the larynx in rats.18 Moreover, anatomical enlargement of the upper pharynx* reflects changes in orientation, alignment and diameters of the basicranium and facial skeleton.19 And Lieberman maintains a suspicion of Heim’s La Chapelle reconstruction, the very specimen on which Lieberman and Crelin established their initial recapitulation idea.
The hyoid bone
The second most prominent anatomical feature of the Neandertal speech debate includes the position of the hyoid bone. The original work done by Lieberman and Crelin1 postulates a rather high position for the missing hyoid bone (and therefore a higher larynx) in the neck, based upon similarities of the basicranium and mandible* of La Chapelle with human newborns and chimpanzees. The hyoid is an important part of the upper respiratory tract and is attached to the larynx to aid as a mediator of muscular forces against it. Its position is of critical concern in determining the effects of its placement upon deglutition (swallowing) and speech.
Morris20 was the first to challenge the high hyoid position for La Chapelle proffered by Lieberman and Crelin, by demonstrating that the angle of the styloid processes* on the skull were inconsistent with their proposed hyoid position. Such a high position, Morris argues, would have made La Chapelle unable to swallow! Since the styloid processes and the greater cornua* of the hyoid are connected to each other by the stylohyoid ligament in a fairly straight line, Morris’ lowering of it makes its estimated anatomical position like that of modern humans.
In the mid-1980s, the only extant Neandertal hyoid bone was discovered at the Mousterian cave site of Kebara in Israel.21 With this discovery came renewed speculation regarding Neandertal speech. A study of the specimen was undertaken to see if any inferences could be made about Kebara’s speech ability.22 Arensburg et al. argue that the morphology* of this hyoid is modern and that the articulatory ability of Neandertal was not impeded as Lieberman and Crelin maintain. In addition, they argue that the hyoid bone and features of the mandible are better indicators of soft tissue placement than is basicranial flexion. This is a reasonable conclusion, because the mandible and hyoid are more closely associated to each other functionally and anatomically than either is to the basicranium. Lieberman, however, argues that the Kebara hyoid is not exactly like that of modern humans in form and cites the inability of Arensburg’s team to statistically discriminate it from that of other animals, including pigs.22 He further states that because hyoid bone morphology is highly variable, it is unlikely that one can reliably reconstruct a vocal tract using it alone.
Research criticisms, limitations and considerations
Other concerns have been raised regarding the research of Lieberman, Crelin and their associates. One is Lieberman’s reconstruction of La Chapelle’s vocal tract while the cranium does not approximate the Frankfort Horizontal.20,23 The Frankfort Horizontal places a human skull in a level position it approximated in life when the subject stands erect and watches the horizon. As one may imagine, the horizontal position of the skull can affect the position of other soft and hard tissue structures in the neck, although this does not worry Lieberman.24 In reconstructing actual soft tissue features of the vocal tract in clay or plastic, as Lieberman and Crelin have done, we begin to move away from the most accurate and conservative arguments we may make about Neandertal anatomy. One case in point, is a reference to Crelin’s4 reconstruction of the Steinheim Neandertal skull as, ‘a piling up of reconstruction upon reconstruction and [a] subsequent abstraction of the argument from the actual fossil remains’,25 an observation found ‘to be most unsettling in considering Lieberman’s arguments’. While Lieberman doesn’t claim to have all the answers about Neandertal speech anatomy, he thinks he and his colleagues have arrived at a workable theory of hominid speech evolution.17
Nearly all of the above criticisms levelled at Lieberman and Crelin question the reliability of soft tissue reconstruction of the vocal tract using osseous* landmarks alone. The human supralaryngeal vocal tract is a complex assemblage of muscle, bone, cartilage and mucosa, not leaving many easily readable traces of itself in bone. As we have seen, scientists studying the phylogenetic* function of this anatomical complex clearly run into problems mainly due to three reasons:
- Historically, reconstructions have been attempted using only one bone at a time.
- Since no Neandertal soft tissue has so far survived for us to study, anthropologists are bound to use modern human populations as a baseline from which to predict archaic soft tissue placement, a proposition that invariably leads to questions of circularity in research.
- The inferences one can make about Neandertal speech ability are limited. While phylogenetic issues can be addressed, ontogenetic and recapitulation ideas like those of Lieberman and Crelin tend to go far beyond what can be reliably deduced from the data.
Lieberman has modified his views slightly since his original research to say that, ‘it is clear that the Neanderthal vocal tract was not, as Edmund Crelin and I thought in 1971, exactly like a human newborn’s. Preliminary data indicate that the vocal tract of a classic Neanderthal fossil like La Chapelle-Aux-Saints probably had the capabilities of at least a two-year-old human child … ’.26 But what Lieberman fails to realize is that Neandertal speech capability was that of an adult Neandertal, not that of any child at all. Apparently, Lieberman is content to hold to his original recapitulation idea, evidence to the contrary notwithstanding.
Budil11 is the first to apply vocal tract features of living people to the problem using measurements that cover the whole supralaryngeal tract. He created a small database of mean* distances for several vocal tract points from computerized tomographs* of 45 adult males to use as a baseline for comparison of the Petralona (a proposed pre-Neandertal) skull from Greece. Budil intended to predict the position of the non-surviving hyoid bone for Petralona using seven landmarks of the cranial floor and mandible. His predicted hyoid position falls easily within the range of modern humans.
Budil’s creation of a database for research using actual living humans instead of cadavers and for incorporating several different types of measurements in his study is praiseworthy. However, there are potential problems with Budil’s work:
- Computerized tomographs, or ‘CT scans’, record radiographic ‘slices’ of the head and axial skeleton in an oblique transverse plane. While the machine is very good at calculating sagittal* dimensions using these slices, it cannot slice directly at the tips of the landmarks Budil studied, unless the scanner is set to a very thin interval. Budil does not say how he measured the tomographs.
- When a patient lies on the CT table, the neck becomes kaiphotic*. This changes the actual positions of soft tissue features within the neck, such as the tongue and larynx, and distorts the ‘at rest’ orientation of the hyoid and mandible.
- In my opinion, Budil facilitated a gross abstraction of the data when he ‘complemented’ the Petralona skull with the Mauer Mandible from Germany in order to obtain mandibular distances, since Petralona’s mandible is missing. His justification for this is that the two are of the same taxonomic affinity and that the Mauer Mandible ‘fits well together with the [Petralona] basicranium’. This is a big ‘no-no’ in anthropology. Mandibles and crania (a la Piltdown) should not be mixed.

Click here for larger view
There are several ways one can study the vocal tract, and none of them are without limitations. The above research history demonstrates that comparisons of archaic human speech anatomy to modern humans using hard tissue landmarks alone is problematic. Perhaps incorporating as many bony landmarks as possible into a study will yield a more accurate analysis.
One study of osteological relevance to phonemic speech has not been addressed: the relationship of the cervical vertebrae to the supralaryngeal tract. In the case of La Chapelle, a point has been made that ‘one cannot locate the [hyoid] bone relative to [its] cervical vertebrae, since the crucial second, third and fourth cervical verterbrae are missing in this specimen’.27 This is a curious statement, since no landmarks exist on cervical vertebrae themselves to suggest relative hyoid position anyway. But is it possible that a fully articulated cervical vertebral column can indicate hyoid position and height of the larynx in the Neandertal throat?
It is logical to conclude that the pharynx and larynx are under greater developmental and selective constraint by the human respiratory complex and the skull than by the vertebral column, since these are more functionally integrated. Therefore, they are better indicators of hyoid position than the cervicals. But it is not unreasonable to suppose that the cervicals can predict the vertical height of the hyoid (and larynx) in the tract. We know from cadaver, radiograph and video fluoroscopic studies, that they occupy a rather restricted envelope of space in the throat, somewhere at a level between the third and fifth cervicals.28 However, trying to determine the vertical placement of the hyoid by guessing the specific positions of each vertebra in La Chapelle is too subjective an approach due to the high variability in hyoid height in modern populations. Judging hyoid height based upon total vocal tract length is less subjective and easier to do.
Hyoid position may be predicted from the length of the neck, using the surviving La Chapelle cervical vertebrae, assuming that soft tissue placement in archaic humans does not differ significantly from that of living modern humans. This work represents a study of modern human vertebrae and basicrania used as a baseline to predict the relative placement of the hyoid bone and epiglottis* in three archaic humans, La Chapelle, Skhúl V and Shanidar-1. This study tests the Lieberman and Crelin argument that a flat basicranium means a higher hyoid placement. It also indirectly tests the limit of osteology’s* contribution to soft tissue reconstruction. Admittedly, it is a circular approach to human phylogenetic research, yet one hopes it will shed some light upon the ‘humanness’ question surrounding Neandertals and the variability of this complex anatomical region.
Materials and methods

Click here for larger view
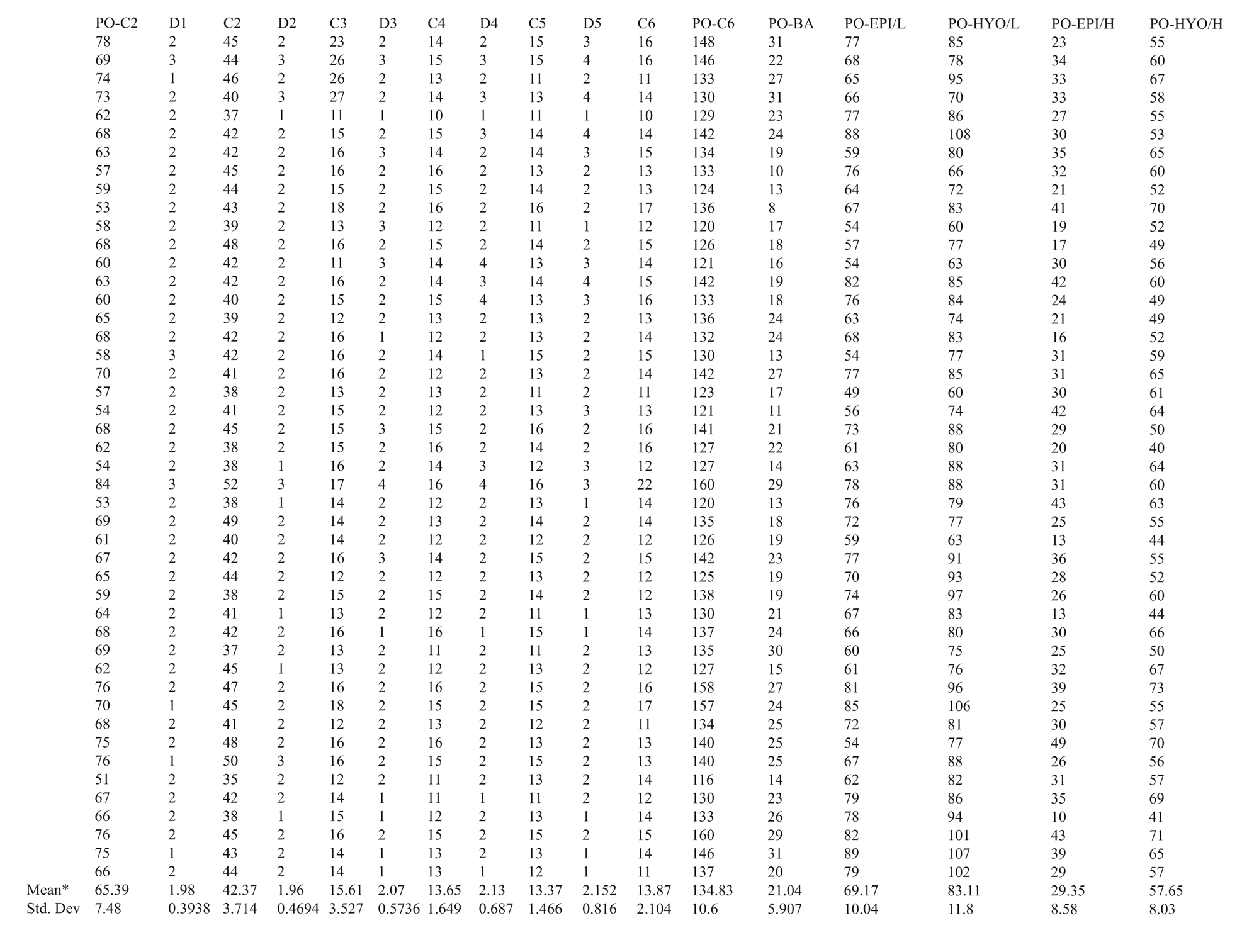
This study consists of three sampled groups, one archaic and two modern. The first modern set was a radiograph sampling of 46 males aged 24 to 65 (since the available archaic fossils studied are also adult males) and of mixed racial affinity from the University of New Mexico, Albuquerque Medical Imaging Center (Table 1). All radiograph measurements were taken in the mid-sagittal plane, either perpendicular or parallel to the Frankfort Horizontal to eliminate concerns of vocal tract distortion associated with atypical head position. Only radiographs of non-pathological, free-standing individuals were used. Correction for x-ray magnification (parallax) was made under consultation with radiologists and by comparison with an actual skeletal sample (see below). An overall magnification error of 18 % was noted and corrected for. Measurements were checked three times for accuracy and repeatability on 20 of the radiographs. This sample served to determine soft tissue placement patterns of the larynx to be used as a baseline for comparison with the archaic specimens.
The second modern sample, consisting of 26 ‘normal’ adult males of mixed racial affinity, was drawn from the Maxwell Museum of Anthropology documented skeletal collection at the University of New Mexico, Albuquerque (Table 2). Age, sex and race were all confirmed by documentation. This sample was used as a comparison and correction gauge to the radiograph sample. Ten of the skeletal specimens were checked three times for accuracy. Obviously, only osseous (bony) tissues could be measured from this sample, but it was valuable in correcting the radiograph sample data.
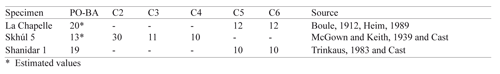
Finally, a sample of three adult male archaic humans was taken (Table 3). This included surviving cervical vertebrae and crania from La Chapelle, Skhúl V and Shanidar-1. A description of each archaic specimen follows.
The La Chapelle Neandertal was discovered in a cave near the village of La Chapelle-Aux-Saints, France in 1908. The cranium is mostly complete and three of the surviving vertebrae are cervicals (C5–C7), two of which are used in this study. Because a recent reconstruction of the La Chapelle skull has resulted in a change to the basicranium,14 an estimate was made of porion*-basion height at 20 mm as derived from the new published measurements.
Click here for larger view
Skhúl V is an adult male hominid of robust modern features. His remains, and those of other moderns, were found within a cave at Mount Carmel, just Southeast of Haifa, Israel.29 Age estimates place these specimens as contemporaries with the classic Neandertals of Europe.30 Most of the cranial vault has survived, but some of the splanchnocranium (face) and a portion of the cranial base is missing. Therefore, his Porion-Basion height is estimated at 13 mm. Three cervical vertebrae (C2–C4) survive, all of which are used in this study.
The Shanidar Neandertals represent some of the easternmost Neandertal finds. A total of nine individuals were found buried in a cave in the Shanidar Valley of Iraq.31 The ‘Old Man,’ Shanidar-1 was discovered in 1957. Except for a missing portion of the left parietal bone, his skull is in relatively good shape, pathologies notwithstanding. Cervical vertebrae five, six, and seven all survive and C5 and C6 are used in this study.
Since radiograph measurements were collected as coordinate data using porion as a datum, it was possible to map the placement of the hyoid, epiglottis, and bottom of C6 relative to porion to actual scale on graph paper. Coarse observation of the radiograph sample was also made using computer scatterplots to check for non-linearity. Means and standard deviations for each radiograph measurement were noted (Table 1). Coefficients of variation (CV = 100 x standard deviation/mean) were also calculated for each set of measurements in order to assess the amount of intrinsic variation. CVs suggest the reliability of the data in demonstrating useable patterns, and are a necessary step before regression formulae are created.

Click here for larger view
Linear multiple regression formulae were obtained from the radiograph sample in order to predict the non-surviving features of each archaic vocal tact (Table 4). Efficiency test values were run for each. The Skhúl V porion-epiglottis length regression formula is the only one to exceed the P = 0.05 efficiency test*, but because no other possible combinations would yield a better alternate, this prediction formula was made to stand.
Two hypotheses were addressed in this study. Hypothesis One asks whether the archaic vocal tract length is the same absolute size as modern humans. One can assume it is not, due to over-all stature differences between moderns and archaics. So Hypothesis Two asks whether the relative placement of archaic features in the vocal tract is the same as in moderns. If the relative distances are shorter than those for modern humans, then the data would tend to support the Lieberman-Crelin model for a high larynx and impeded speech capability.
Click here for larger view
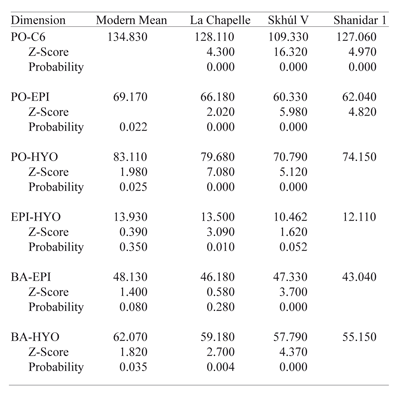
Six (absolute) vertical distance predictions were derived for each archaic specimen and Z-tested* against the modern radiograph sample means of identical measures (Table 5). Next, predicted archaic vertical distances were converted to indices by dividing each distance by the total tract height (PO–C6), as was done for the modern sample means (Table 6). Index values for the vertical dimensions display placement of each feature relative to the total tract length, and place both the modern and archaic data on the same quantitative level for comparison.
Results
Absolute and relative distance values were calculated using regression formulae results derived from a modern human sample as noted above (Tables 5 and 6). CVs among the modern human variables were calculated also (Table 7). CVs demonstrate which measurement groups contain the greatest amount of intrinsic variation. The EPI-HYO measurement (see Figure 1) is the most variable. This high degree of variability colours the interpretive strength of the archaic sample data for the same measurement, since the measurement is partly derived from the modern sample data. And therein lies the problem with circularity in this kind of research. But it also implies that the distance between the hyoid bone and the tip of the epiglottis is not as critical to the overall function of the vocal apparatus as how far either location is from the cranial base. This variation can be clearly seen with the naked eye on the modern sample radiographs, and one may presume that it does not impede or enhance speech.
The modern human radiograph data were scatter-mapped. The predicted values for total tract length, epiglottis vertical displacement and hyoid bone vertical displacement from the archaic regression predictions were sketched into the map for comparison (Figure 2).
Analysis of total tract length, vertical hyoid displacement and vertical epiglottis displacement are as follows. The modern human adult male vocal tract averages 135 mm in length. The modern human epiglottis averages 69 mm below porion, or 51 % of the distance between porion and the bottom of C6. The epiglottis averages 48 mm below the cranial base (basion) and or 61 % of the distance from basion to the bottom of C6. The anterosuperior* hyoid body in modern males rests an average of 83 mm below porion, accounting for 62 % of the distance from porion to C6, and averages 62 mm below basion or 73 % of the distance from basion to C6. The average vertical distance between the tip of the epiglottis and anterosuperior hyoid body is 14 mm, representing 10 % of the distance between porion and C6. Table 5 lists the archaic sample vocal tract predictions and their corresponding Z-scores and probability factors against the means of the same measurements in the modern human sample.

For the most part, actual (absolute) archaic human vocal tract dimensions do not fall within the range of modern humans. This comes as no surprise, since the overall stature of archaic humans is considerably shorter than that of modern humans. The longest total vocal tract length among the archaics is that of La Chapelle (128 mm), who falls beyond four standard deviations from the modern human mean. The more ‘modern’ archaic, Skhúl V total tract length is the shortest of the three at 109 mm. The very short tract length is due in part to the flatness of the cranial base, with a vertical distance from porion to basion of only 13 mm as compared with La Chapelle (20mm) and Shanidar (19 mm). One interesting find is that the EPI-HYO predictions are more like that of modern males in the case of Shanidar and La Chapelle. This suggests that for their overall neck length, these two archaic males had a taller larynx than expected. The comparisons also reveal that the distance from the base of the skull to both the hyoid and epiglottis in both La Chapelle and Skhúl are within the modern human range also, refuting the Lieberman and Crelin argument. Implications of this are explored in terms of relative distances below. Overall, actual dimension comparisons confirm that these two archaic adult males did not have vocal tract dimensions as large as the typical modern human male.
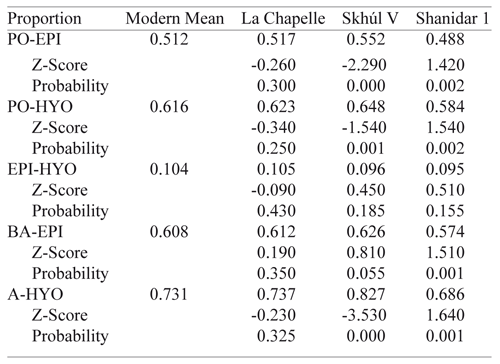
An analysis of the relative placement of vocal tract features was done to test Hypothesis Two (Table 6). Z-scores and probability factors were calculated against the means of identical measurements in the modern sample. Comparisons demonstrate that the La Chapelle Neandertal exhibits a complete resemblance to modern humans in neck morphology; none of his relative dimensions fall outside of the modern human range. For his size, La Chapelle’s neck anatomy was not significantly different from that of modern humans. The comparisons of the Skhúl and Shanidar specimens differ, however. The Skhúl V neck morphology matched modern humans in only one respect; the vertical distance between the epiglottis and hyoid. In all other respects, Skhúl V is significantly different from the modern human mean values, yet not in the direction that Lieberman, Crelin and their advocates would like. It appears that this more ‘modern’ specimen’s larynx was much lower in the throat than in modern humans (10 % lower), despite the fact that Skhúl V’s cranium is the flattest of the group. Conversely, the Shanidar-1 epiglottis and hyoid are significantly higher in the neck than in moderns. This finding argues against the Lieberman and Crelin model, yet in the opposite direction. Since the Shanidar cranial base is fairly high (19 mm) one would expect, according to the model, that the larynx would be lower in the throat. The vertical distance between the epiglottis and hyoid in all three archaic specimens is within the range of modern humans, although the CV for the modern sample (54.77) is very high and reduces the reliability of the data, as mentioned above. The resulting predicted relative distances accept Null Hypothesis Two, eight times out of eighteen measurements, with five of those predictions belonging to one individual, La Chapelle. The results of this study, while refuting Lieberman and Crelin, do so in a totally unexpected way. While the Shanidar Neandertal has a higher predicted larynx position than moderns, it also has a high basicranium. Yet, Skhúl V, of modern human-like, robust skull morphology, doesn’t share a modern human vocal tract with us. This is a curious discovery owing to the fact that this individual shares features of both modern and Neandertal morphology and is a Neandertal contemporary. Finally, the La Chapelle Neandertal shares a fully modern vocal tract morphology with us. The basicranial height measurement for La Chapelle (20 mm) was estimated using other cranial measurements from the Heim reconstruction.14 The original Boule reconstruction for this specimen32 reveals a basicranial height of 18 mm. There is not enough difference in these two measurements to render a significantly different vocal tract prediction. But in each of the three cases, the Lieberman-Crelin model, that a flatter cranial base means a higher larynx and impeded speech, is discredited.

Providing that soft tissue anatomy, motor control and cognitive linguistic ability were not significantly different from modern humans, the La Chapelle and Skhúl V archaics probably enjoyed the same range of vowel sounds that we do today. From the data, however, it is impossible to tell if the Shanidar-1 Neandertal did. Such a high predicted larynx position may have impeded vowel sounds or possibly the ability to swallow!
Analysis, observations and research considerations
Certainly, there exists a fairly high degree of variability in vocal tract anatomy among humans, and especially among archaic humans. While we know that modern human variation in many skeletal traits is already high, among archaic humans it is even greater. Thus we may wish to examine the hypothesis that after the Flood an event or series of events have occurred to reduce human variability over time to the present day.
The results of both the absolute and relative values raise the possibility of allometric* differences. The relative position of vocal tract features is a function of the absolute size and shape of the Neandertal and modern human vocal tracts, respectively. Allometric studies may shed more light upon the Neandertal speech question and upon our relatedness to Neandertals in general and would be an excellent study for any creation scientist wishing to conduct research on Neandertal relatedeness to modern humans.
Reconstruction of the archaic human vocal tract is hampered by the limited amount of data one can collect from incomplete skeletal remains. So, what can really be known of soft tissue anatomy using only osteological data in the genus Homo? Probably the most important result of this study is that it demonstrates the difficulty in inferring soft tissue placement from skeletal evidence alone. We may wish to be very careful anytime we argue soft-tissue anatomy using only hard tissue remains. Even the casual reader in creation science is well aware of the pitfalls associated with attempts at soft tissue reconstructions and the ways in which they have shaped popular evolutionary thought. The same pitfalls await any imprudent, yet zealous creationist wishing to make a point.
Finally, it should be understood that while any experiment may or may not tend to support the Lieberman-Crelin argument for limited linguistic ability, these results can only indirectly address the relatedness of Neandertals to ourselves. As Grover Krantz puts it, ‘the lack of full speech capability in Neandertals does not in any way remove them from our ancestry, as some have claimed’.33 Keeping things in perspective, it is important to remember that there are human languages on earth today that do not make use of ‘high’ vowel sounds, yet it does not impede the speaker’s ability to communicate intelligently or effectively. Overall, these findings tend to support the work of Morris,20 Houghton23 and Arensburg, et al.,22 who have maintained that Neandertals had a modern-placed hyoid and were probably capable of articulate speech. But even if such were not the case, it does not necessarily follow they are non-human. More creationist anthropological field and laboratory studies should bear this out.
Acknowledgements
I am indebted to James Dawson for statistical advice and also to Erik Trinkaus, Kurt Wise and Linda Walkup for their counsel. Thanks also to the Maxwell Museum of Anthropology at the University of New Mexico for access to its skeletal collection and to UNM Medical Center Imaging Library.
Glossary |
|
| Allometric | the growth of body parts at different rates |
| Anterosuperior | upper front |
| Basicranium | base of the skull |
| Computerized tomograph | precise type of x-ray imaging that can show ‘slices’ of a specimen at given intervals |
| Cranium | skull |
| Efficiency test | a mathematical (statistical) test for determining the reliability of a statistical result. P=0.05 means that 95 times out of 100 trials the same result would occur and suggests that the data is reliable. |
| Epiglottis | a flap of cartilage at the root of the tongue, which is depressed during swallowing to cover the opening of the windpipe. |
| Greater cornua of the hyoid | tips of the horseshoe-shaped hyoid where ligaments attach toward the base of the skull |
| Hyoid | horseshoe-shaped bone that surrounds the voicebox and creates the ‘Adam’s Apple’ |
| Kaiphotic | straightening of the naturally curved vertebral column |
| Larynx, larynges | voice box |
| Mandible | lower jawbone, consisting of one piece in humans and most apes |
| Mean | statistical average |
| Morphology | shape or physical characteristics |
| Ontogenetic | development of an organism from fertilization to adulthood |
| Osseous/osteology | bony or having to do with bone |
| Pharynx | throat area behind the nasal cavity and mouth |
| Phylogenetic | evolutionary/developmental history of organisms |
| Porion | name for a landmark locating the uppermost margin of the ear-hole in the side of the skull |
| Recapitulation theory | an evolutionary idea that says that development of an organism toward adulthood recapitulates or mimics its evolutionary history |
| Sagittal | referring to the mid-plane of the body, front to back |
| Splanchnocranium | facial skeletal complex |
| Styloid process | bony landmark at the base of the skull, serving as an attachment for a ligament that attaches to the hyoid bone in the throat |
| Z-test | simple comparison of sample data with a normal distribution (bell curve) |
Related products
References
- Lieberman, P. and Crelin, E.S., On the speech of Neanderthal man, Linguistic Inquiry 2:203–222, 1971. Return to Text.
- Laitman, J.T., Heimbuch, R.C. and Crelin, E.S., The basicranium of fossil hominids as an indicator of their upper respiratory systems, Am. J. Phys. Anthrop. 51:15–34, 1979. Return to Text.
- Lieberman, P., On the Origins of Language: An Introduction to the Evolution of Human Speech, Macmillan, New York, 1975. Return to Text.
- Crelin, E.S., The Steinheim skull: a linguistic link, Yale Scientific 48:10–14, 1973. Return to Text.
- Lieberman, P., Crelin, E.S. and Klatt, D.H., Phonetic ability and related anatomy of the newborn and adult human, Neanderthal man and chimpanzee, Am. Anthrop. 74(3):287–307, 1972. Return to Text.
- Laitman, J.T., Heimbuch, R.C. and Crelin, E.S., Developmental changes in a basicranial line and its relationship to the upper respiratory system in living primates, Am. J. Anat. 152:467–482, 1978. Return to Text.
- Houghton, P., Neandertal supralaryngeal vocal tract, Am. J. Phys. Anthrop. 90:139–146, 1993. Return to Text.
- Frayer, D., Language capacity in European Neanderthals (in Press). Return to Text.
- Burr, D., Further evidence concerning speech in Neandertal man, Man 11:104–110, 1976. Return to Text.
- Gibson, K.R. and Jessee, S.A., Cranial base shape and laryngeal position: implications for Neanderthal language debates, AAPA Meeting, 1993. Return to Text.
- Budil, I., A functional reconstruction of the supralaryngeal vocal tract of the fossil hominid from Petralona; in: Studies in Language Origins Vol. 3, John Benjamins, Philadelphia, 1994. Return to Text.
- Hiiemae, K. and Palmer, J.B., Tongue, palate, hyoid and the capacity for speech, AAPA Meeting, 1993. Return to Text.
- Le May, M., The language capability of Neanderthal man, Am. J. Phys. Anthrop. 42:914, 1975. Return to Text.
- Heim, J.L., La nouvelle reconstitution du crane Néandertalien de La Chapelle-Aux-Saints méthode et résultats, Bull. et Mem. de la Soc. d’Anthrop. de Paris 1(1–2):95–118, 1989. Return to Text.
- Frayer, D., Evolution at the European edge: Neanderthal and Upper Paleolithic relationships, Préhistoire Européenne Périodique 2:8–69, 1992. Return to Text.
- Heim, J.L., Les hommes fossils de La Ferrasie (Dorgone) et la problème de la definition de Neandertaliens classiques, L’Anthropologie 78:376–377, 1974. Return to Text.
- Lieberman, P., Hyoid bone position and speech: reply to Dr Arensburg, et al. (1990), Am. J. Phys. Anthrop. 94:275–278), 1994. Return to Text.
- Laitman, J.T. and Reidenberg, J.S., Advances in understanding the relationship between the skull base and larynx with comments on the origins of speech, Human Evolution 3:99–109, 1988. Return to Text.
- Bosma, J.F. and Fletcher, S.G., The upper pharynx, a review, Part 1: Embryology and Anatomy, Annls. Otol. Rhinol. Laryngol. 70:53–73, 1961. Return to Text.
- Morris. D.H., Neanderthal speech, Linguistic Inquiry 2:15–20, 1974. Return to Text.
- Bar-Yosef, O., Vanderneersch, B., Arensburg, B., Goldberg, P., Laville, H., Meignen, L., Rak, Y., Tchernov, E. and Tillier, A.M., New data on the origin of modern man in the Levant, Current Anthrop. 27:63–64, 1986. Return to Text.
- Arensburg, B., Schepartz, L.A., Tillier, A.M., Vanderneersch, B. and Rak, Y., A reappraisal of the anatomical basis for speech in Middle Paleolithic Homonids, Am. J. Phys. Anthrop. 83:137–146, 1990. Return to Text.
- Houghton, P., Neandertal supralaryngeal vocal tract, Am. J. Phys. Anthrop. 90:139–146, 1993. Return to Text.
- Lieberman, P., On the Kebara KMH 2 Hyoid and Neanderthal speech, Current Anthrop. 34(2):172–175, 1993. Return to Text.
- Carlisle, R.C. and Siegel, M.I., Additional comments on problems in the interpretation of Neanderthal speech capabilities, Am. Anthrop. 80:367–372, 1978. Return to Text.
- Lieberman, P., Eve Spoke, W.W. Norton & Company, New York, p. 94, 1998. Return to Text.
- Falk, D., Comparative anatomy of the larynx in man and the chimpanzee: implications for language in Neanderthal, Am. J. Phys. Anthrop. 43:123–142, 1975. Return to Text.
- Williams, P.L., Warwick, R., Dyson, M. and Bannister L.H., Gray’s Anatomy, 37th Ed., Livingstone, New York, 1989. Return to Text.
- McGown, and Keith, A., The Stone Age of Mt. Carmel, Clarendon Press, 1939. Return to Text.
- Klein, R.G., The Human Career, U. of Chicago Press, 1989. Return to Text.
- Trinkaus, E., The Shanidar Neandertals, Academic Press, London, 1983. Return to Text.
- Boule, M., Annales de Paleontologie, Tome VII, Masson E.T., cie Editeurs, Paris, 1912. Return to Text.
- Krantz, G.S., Sapienization and speech, Current Anthropology 21(6):773–779, 1980. Return to Text.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Readers’ comments
Comments are automatically closed 14 days after publication.