

The world of bats

Marvels of design
There are hundreds of species of bats (chiroptera) in the world. And they are able to colonise a wide variety of habitats, utilising food sources ranging from fruit and pollen to fish and insects.
Bats deserve careful study, as they exhibit features not found in other mammals. The most important of these is the power of flight, using a stretched membrane which acts as an aerofoil (wing).
Interest in bats is widespread for a number of reasons. Conservation is now vital for some species if they are to survive; vandalism is easy; and colonies in caves can be wiped out in minutes. Chemical insecticides can also take a heavy toll on bats. Biologists cannot forget that DDT has been found in so many forms of life, nor can they minimise the serious effects of this compound on fertility in birds and bats, especially where they are at the top of the ecological pyramid of predation.
Serious interest in bat welfare goes back to the American bat towers erected by Dr Charles Campbell1 in Texas from 1908 to 1911, and it was Texas which passed the first Bat Protection Law in 1917.
Common characters
Bats share a considerable range of characteristics common to most mammals: they secrete milk, all have hair to a lesser or greater extent on the skin, and the lower jaw is composed of a single bone (dentary), whereas birds and reptiles have a complex of bones. In the ear, bats have three bones (ossicles), and their milk teeth are replaced in part by one more set. They are able to maintain a steady body temperature during the active phase of the annual life cycle, while allowing it to drop during hibernation. Before birth, young bats develop in the uterus, being nourished by the typical mammalian structure called the placenta, which grows from the uterine wall.
Unique bat features
Only the chiroptera, among all the mammalian orders, possess wings and employ echo-location for food capture. The Greek name of ‘hand wing’ indicates that the five radial finger bones are used to stretch the skin between the fore and hind limbs; a few species have an additional membrane of skin, linking the hind limbs and the tail vertebrae. The Creator’s design for this remarkable wing also involves a special muscle running along the leading edge of the wing which, when it contracts, causes a change in the camber of the wing, thereby affecting its aerodynamic potential. The wing membranes enclose blood vessels which transport nutrients to the flight muscles and also act as heat-dissipators when cooling is necessary. In some bats, the expanse of the wing allows carbon dioxide to diffuse from the veins, thus acting as an excretory surface.
The unique feature of echo-location was suspected by Italian scientist Lazzaro Spallanzani (1729-1799) after he had performed experiments with bats in darkened rooms. Later, Swiss naturalist Charles Jurine concluded that bats could capture prey through an ability to hear reflected sounds.
Hiram Maxim was the one who considered the possibility of a sonic warning device in bats, although he wrongly suggested that the frequency was very low and was generated by the wings. Modern investigations started in the late 1930s. Professor G.W. Pierce developed an ultrasonic detector with the help of Donald Griffen. Captain Bernard Acworth, the founder of the Creation Science Movement, was also involved in the development of a sonar device to be used against submarines.
We now know that bats use oral emitters (their mouths) to produce a series of short pulses of high frequency; a few species use their nostrils as emitters. When this sound strikes an object, it is reflected as an echo.
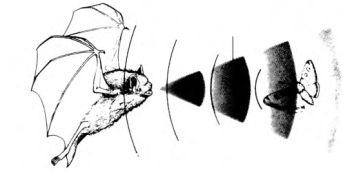
The sound frequency range used by bats varies from species to species, with the majority using high frequencies between 20 and 80 kilohertz. A few use 4-5 kHz and very exceptionally others go as high as 120-210 kHz. Design in this sphere appears to involve a careful correlation between shortwave sounds and the average size of the prey being in the region of 5-20 millimetres in diameter.
To further complicate the issue (and preclude the possibility of small evolutionary changes being effective for survival), many bats have a repertoire of signal types depending upon their immediate use, location and density of vegetation. This demonstrates the remarkable flexibility built into their design by the Creator. Many fruit-eating species do not echo-locate, flying by day or at dusk and finding their food by normal sight among the tropical fruit trees.
To seal the utter impossibility of evolutionary development, we also know that certain moth families (the noctuidae and geometridae, for example) contain species which are able to detect marauding bats and take evasive action to avoid predation. The wonder of the situation is even more enhanced by the presence in some moths of an organ capable of generating a ‘jamming’ signal which disturbs the bat’s echo patterns and confuses its hunting technique! Research has been undertaken to determine the extent to which a bat can modify its own signals to avoid this ‘jamming’ by the insect prey; such variation renders one almost speechless in praise of this superb functional ability, which is the hallmark of Divine wisdom and foreknowledge.
It will be realised that the echo-location apparatus must be perfect to be of any use. Both emitter and receiver must co-operate together, besides being coupled to the mental computer in the animal’s brain.
Reproduction
In common with other placental mammals, the chiroptera undergo a prenatal period (gestation), develop from a single fertilised egg, and are born in a highly developed condition. This enables them to feed instantly on their mother’s milk even in flight, using their hooked milk teeth for a good grip. Usually only one batling is produced per oestrous cycle; twins and triplets are relatively rare.
One peculiar feature sometimes found in bats living in cooler habitats is delayed ovulation. This is linked with over-winter sperm storage and coincides with prolonged hibernation among many north temperate species. Batlings born in late spring or early summer find abundant insect life in northern Europe and the British Isles. The period of gestation varies from 40-60 days in the small microchiropterans (insect eaters) up to eight months in the tropical vampires (megachiropterans).
Birth itself often involves the mother’s helping her offspring out of the birth canal by gripping the tiny batling with her teeth and tugging; bats with a tail membrane use this as a cradle after delivery. The females with young form a unisexual nursery colony from which males are excluded. The nursery breaks up when the development is sufficiently advanced for the young to forage for themselves, and then the females rejoin their partners for the remainder of the flying season.
Fossil bats

Superb fossil bat specimens are available from all over the world in many of the higher, or ‘younger’ strata of the so-called geological column. This is to be expected, since many could have avoided Noah’s Flood for some time, succumbing only when all their perches had disappeared below the waves. A rapidly killed specimen from the so-called ‘Middle Eocene’ strata of Grube Messel in West Germany even shows undigested moths in its stomach.
There are no intermediate fossil forms between bats and something else, nor do any proto-bats exist. Bats’ features are so precise and specific that partially developed individuals would be incapable of surviving in the wild. Bats appear in the ‘fossil record’ fully developed, fully functional, and fully recognisable only as bats. Their modern-day counterparts, therefore, are merely products of reproduction ‘after their kind’.
Conclusion
When we consider the amazing design features in bats, such as the complicated echo-location system which would be of no use in ‘partially evolved’ stages, and we note the total lack of fossil evidence for the ‘evolution’ of bats, we must conclude that there is no evidence for the evolution of bats. Their function and design can only point to their creation in the beginning by God Himself.
References and notes
- Jennings, F.W., Dr. Charles Campbell and San Antonio’s Bats, uiw.edu. Return to text.










Readers’ comments
Comments are automatically closed 14 days after publication.