

Journal of Creation 13(1):115–125, April 1999
Browse our latest digital issue Subscribe
Did God make pathogenic viruses?
Summary
A review of the structure, function, and role of viruses in ecology is presented. It is concluded that viruses are non-living entities, similar to seeds and spores whose functions include carrying genes from one plant or animal to another. Viruses are a part of a system that helps to produce the variety that is critical for life and, importantly, they carry resistance to disease from one organism to another. Most viruses live in their host without causing problems. Pathogenesis is evidence of something gone wrong, a mutation or the accidental movement of genes, and not evidence of a system deliberately designed to cause human disease and suffering.
Introduction
A major line of reasoning used to argue against the creationist worldview is ‘why a benevolent God would create pathogenic organisms whose sole function seems to be to cause disease and suffering?’ The account of Noah and the flood is often criticised by the claim that God must have wanted pathogenic viruses in the world: because they exist today, God must have brought them on the ark.1 One evolutionist summarised this view as follows:

‘ Although there is enormous beauty to be found in the plants and animals of this world, as well as in the physical planet itself, there is also much ugliness. Beauty and ugliness are, of course, human perceptions, but then so is the belief that an intelligent Creator is necessary to explain the existence and nature of the world. Innumerable examples could be cited of rather nasty biological realities which are perfectly understandable in terms of evolution, but which make no sense whatever in terms of design by an infinitely intelligent, wise, and compassionate Creator. Every living creature, including plants, must contend with the ravages of diseases and parasites. Most disease-causing bacteria and viruses, which exist in encyclopedic profusion, serve no useful “purpose” whatever except to infect other creatures and to make their lives more difficult or shorter.’2
The word ‘virus’ elicits memories of virus plagues such as the 1918 influenza plague that killed up to an estimated 20 million people.3 The common public perception is that the only function of viruses is to cause disease. Among the many common diseases caused by viruses are cold sores, hepatitis B, herpes, yellow fever, viral meningitis, chicken pox, colds, mononucleosis, mumps, rabies, polio, shingles, smallpox, warts, viral pneumonia, AIDS and some cancers.
Viruses can also produce health problems by influencing the immune system to attack the body, resulting in auto–immune diseases such as diabetes, lupus erythematosus, multiple sclerosis, and rheumatoid arthritis. Viruses can cause auto–immune diseases by leaving parts of their DNA in their host which may cause their protein–immune fingerprint to become embedded in the hosts’ cell membranes. When these cells reproduce, the daughter cells also possess these unique markings. White blood cells may mislabel these self-cells as foreign and inappropriately attack them. In Hepatitis B infections, more liver damage may be caused by the immune system’s attack on the infested liver cells than by the actual virus. Viruses have even been implicated in causing some cancers such as leukaemia and are also a scourge of farmers, infecting both livestock and their crops.
Although viruses were discovered only at the turn of this century, research has now found a substantial amount of evidence that they serve several major roles in ecology and are actually essential for life. Without viruses, the genetic revolution we are now experiencing would be impossible. They also serve numerous beneficial functions that we are just beginning to research and understand.
The discovery of viruses
Scientists first discovered and studied this mysterious form of ‘life’ at the turn of the century. Researchers found that if the fluid extract from certain diseased animals and plants was dissolved in a solvent and passed through the finest filter known then (unglazed ceramic plates were often used) the filtrate still caused disease.4 Russian Scientist Dmitri Ivanovski found the filtrate from a section of a tobacco mosaic disease infected leaf would infect healthy plants.5
Dutch botanist Martinus Bijerinck called this infectious fluid contagium vivum fluidum. This fluid contained agents that are now known as viruses, the Latin word for ‘poison’. At first, many researchers assumed that toxins must have caused the disease. Bijerinck disproved this theory when he showed that the sap could successively transmit the fully virulent disease through a large number of plant generations. This indicated that the disease agent was multiplying in the plant, otherwise it would have become successively weaker as it spread to each new plant generation. Others speculated that bacterial spores far smaller than those that had been discovered were the cause of the disease. It was eventually realised that the culprit was a non-cellular form of ‘life’ that could diffuse through the cell walls and membranes into the cell’s protoplasm.
William Elford of the National Institute for Medical Research in London developed a new filter technology in 1931 which helped researchers to realise how extremely small viruses are.6 We now know that comparing a virus to an animal cell is like comparing a basketball to the New York World Trade Center. It was only since the 1930s, with the invention of the electron microscope, that scientists could actually visualise viruses.
In the middle 1930s, Wendell Stanley of the Rockefeller Institute mixed viruses in a solvent. He then allowed the solvent to evaporate and discovered that crystals formed. The fact that crystals formed meant that all the virus units have nearly identical shape, weight, charge, and chemical characteristics. Many viruses, including Reoviridae, Parvoviridae and Iridoviridae are regular icosahedrons having exactly 20 triangular faces, 12 vertices and 30 edges (Figure 1).7 This is clear evidence that viruses are unlike any known living thing and are much more like inanimate than animate matter. Some viruses are isometric shaped, others resemble long round tubes, and still others give the appearance of miniature rocket ships. Debate as to whether viruses are living or nonliving ensued soon after Stanley’s discovery. Today we recognise that viruses are gene carriers just as lipoproteins are the cholesterol/fatty acid carriers of the body’s circulatory system.
The construction of viruses
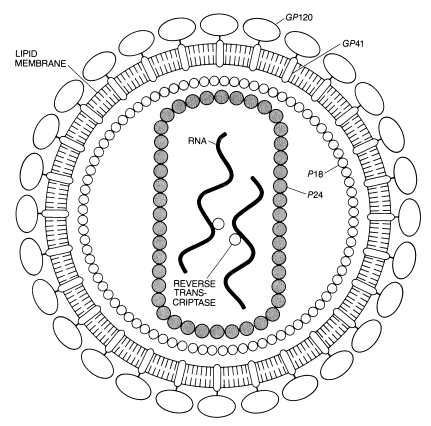
Five basic morphological shapes of viruses exist: spherical, cylindrical, brick, bullet and tailed. Viruses are contained by a protein cover called a capsid that is often coated with an envelope made of carbohydrates or lipids—an icosahedral capsid is made up of 122 capsomers, of which 110 are hexameters and 12 are pentameters.8 This coated capsid houses nucleic acid and other structures that facilitate preserving the genes they contain (Figure 2).9 The nucleic acid may be either double or single stranded DNA or RNA. If RNA is used as the master code, the virus is called a retrovirus and requires the complex enzyme called reverse transcriptase to convert the RNA to DNA. Finally, all viruses contain proteins (called antigens) extending from their surface that can bind with specific receptors on the host cells.
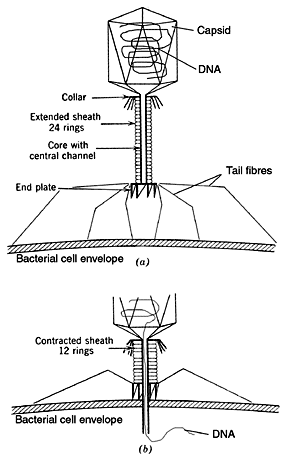
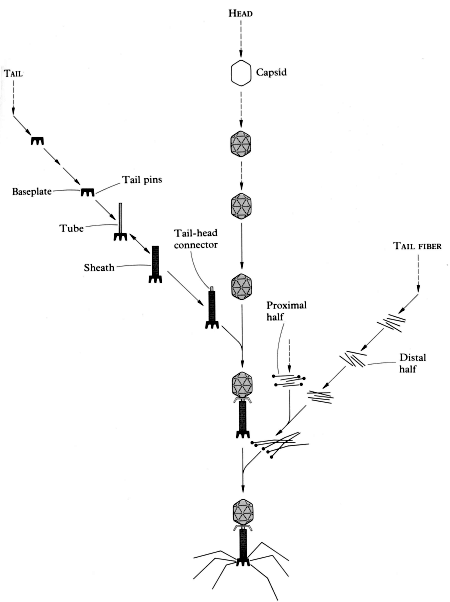
Some of the more complex viruses also have accessory structures that enable them to attach to selected organisms. These structures include a long tube-like sheath, several tail fibres, and an injector (Figure 3).10,11 Each of these seemingly simple structures is enormously complex, and each has hundreds or thousands of parts. The research devoted to understanding them ‘will keep T-4 [a bacterial virus or bacteriophage] morphogenesis at the leading edge of molecular biology well into the next century.’12
Viruses have none of the characteristics of life—they do not grow, they lack cells, and they come only in standard models with few, if any, variations of standard parts. They lack most of the cell enzymes and organelles needed to live, and consequently must exploit their host’s organelles. For this reason, viruses are called obligate intracellular parasites, and are ‘infectious particles’ rather than organisms. The complete infectious unit is called a virion. The few enzymes they possess (such as integrase) are usually related to the mechanisms they use to enter their host cell. They are usually only able to multiply in their specific host, and often in only a specific organ within the host (such as the liver). All members of one viral type are usually almost identical in every way except for the glycoprotein antigens on their protein coat.13 It is this signal that can trigger an immune system response in a host.
Once it was realised that viruses are gene carriers, the next step was to determine how they carried genes to other cells and spliced them into these cells’ DNA. The contents of the gene package that allowed viruses to carry out their role was another research focus. As the role of viruses became better understood researchers began to try to exploit them for human uses. This gave rise to the genetic revolution, including recombinant DNA technology and gene therapy.
Viral replication
Viruses are the smallest infectious agents known and range from 200 nanometers for vaccinia to 20 nanometers for parvovirus. Compared to animal cells, viruses are extremely small—fully 50 million polioviruses can fit into the average human cell.14 A typical bacterium is 1 micron in diameter, while a bacteriophage is one-fortieth of a micron long. The relation between viruses and their hosts is complex, and usually begins when a virus makes contact with a potential host cell. All known life forms can be ‘infected’ by viruses but some life forms appear to be less prone than others, for example, some species of arthropods and gymnosperms are known to be virus hosts.15
Virus multiplication involves six basic steps:
Attachment. Viruses and all animal cells contain projections, typically glycoproteins, that allow a virus and animal cell to make contact and to chemically and mechanically bond if compatibility exists. The protein coat (and lipid envelope, if present) thus must attach to the outer membrane of the host to infect it. To infect a cell, the antigens on the virus surface must fit into the cell’s receptor sites. If the fit is not precise, attachment cannot occur and penetration will not result. The fit is usually species-specific; thus a specific virus type will infect only a specific animal or plant type. However, some virus types such as rabies and influenza have a wide range of hosts.
Penetration. Subsequent to attachment, most viruses are drawn into the cell by the cell membrane ‘closing in’. This process is called endocytosis and it is the same process the cell uses to take in nutrients. There are some virus types although, that can pass directly through the pores in the host’s cell membrane. There are also others such as bacteriophages which remain outside the cell but inject their DNA into the cells (Figure 3).
Transfection. The viral DNA is spliced into a specific site on the host DNA by integrase. The enzyme does this by cutting the circular plasmid DNA, then splicing in the new DNA and repairing the two splice sites (see Figure 5).
Replication and Synthesis. The viral DNA or RNA directs the host cell to produce copies of viral nucleic acids and proteins, including enzymes.
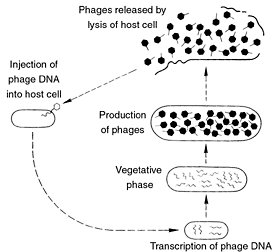
Assembly. Once inside the cell, the virus can set up what might be likened to the biological equivalent of an assembly line (Figure 4)16. In one type of phage the tail is assembled by first constructing the protein scaffolding. Protein building blocks are then added one at a time. This process of addition stops when another protein acting as ‘measuring tape’ determines that the tail is the proper length. A signal is then produced indicating that the structure is complete and the scaffolding protein detaches to be used again in making other virus tails. Herpes and other viruses come with protein tool kits of their own. Most other viruses, such as the tobacco mosaic virus, have to rely almost totally upon the cell’s own tool kits.
Release. The new viruses are released from the cell to infect other cells, spreading even more genes to other cells.17
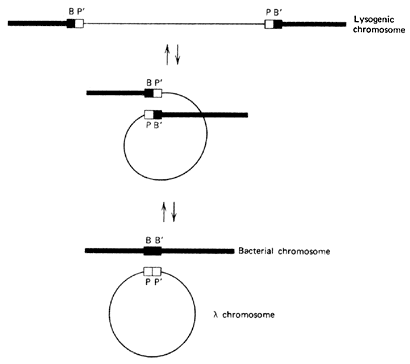
Retroviruses cannot damage cells until they use reverse transcriptase to convert their RNA to DNA. The host then may integrate the virus’ genes in its own DNA, thereby producing a copy of the virus whenever the cell replicates. In this state a phage genome is referred to as a prophage.
The origin of all viruses
Some evolutionists hypothesize that viruses ‘evolved’ from bacteria by natural selection. In this process, as they become parasites, they lost all the complex protein structures that bacteria require. Others hypothesize that viruses were the first form of life, and that bacteria evolved from them (as did all other life). The fatal problem with this theory is that viruses are not living, and in order to reproduce and to make ATP, they require all of the complex cellular machinery present in bacterial cells. Other scientists speculate that a reverse symbiosis occurred, and that viruses arose out of cell parts such as bacterial plasmids and other organelles, and eventually evolved into separate forms of life.18
So far evidence is lacking for each of these theories. Both bacterial plasmids and viruses contain the nucleotide sequences required to initiate replication. While these structures are necessary for the function of each, this does not prove either’s phylogeny. Further, all ‘ancient’ viruses so far discovered in ‘ancient’ amber and other places are fully developed, functional viruses.
The role of viruses in ecology
The importance of viruses is closely related to the importance of bacteria. As Margulis notes, microorganisms have long been considered ‘tiny little beings [that] are primarily germs and pathogens.’19 In contrast to this public image, bacteria are at the basis of our life-support system. They supply our fertile soil and atmospheric gases. They cleanse our water supply, play a role in stabilising the atmospheric nitrogen concentration, regulate the acidity or alkalinity of the soil environment, and thus generally ensure that our world is liveable.20
The view now emerging of the normal relationship between viruses and genes is not so much a host/invader relationship, but a relationship more akin to bees carrying pollen from flower to flower, thus causing cross-fertilisation. Viruses carry not only their own genes, but also those of other creatures as well, especially those of bacteria.21 Although bacteria pass genetic information to each other using several processes such as pili transfer (see below), viral transfer is now known to be critically important.22
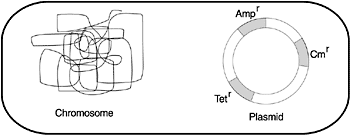
A critical role that viruses play relates to the fact that bacteria contain a constant, stable genetic system (the large replicon), but they function in the world by acquiring and exchanging a diverse set of variable genetic systems (several small replicons, including plasmids, viruses, and so forth). The small replicons are physically separated from the major bacterial DNA, called the genophore. New DNA can be inserted into the genophore; and it usually divides synchronously with it, but some is able to start self-replicating autonomously (Figure 5).
Mathieu and Sonea claim that viruses convert all bacteria into one giant, global ‘superorganism’, and that viruses ‘possess a remarkable mechanism for the creation and exchange of genetic material’.23 A major class of genes exchanged are antibiotic-resistance genes (see figure 6), along with genes that allow bacteria to degrade toxins (such as polychlorinated biphenyls) or convert mercury to less noxious forms.24 This ability is significant in the development of resistance to antibiotics produced by other organisms, allowing bacteria to survive and helping to maintain the balance so critical for ecology.
An important category of small replicons are the prophages, phages and other virus types. Further, no bacterium ‘has yet been found in nature without such a temporary genetic supplement.’25 Some of the replicons, especially those that are not useful to bacteria, eventually disappear. This process was once called curing because the replicons were then considered to be harmful infectious elements. We now know that the role of these viruses is important in conferring variation to bacteria. For this reason, viruses are critical for bacteria, and bacteria are critical for ecology.26To produce this variation
‘each small replicon may visit thousands of different bacterial strains, and each bacterial cell, although usually harboring only a few different small replicons at a time, is able to be visited by tens or hundreds of different types.’27
Bacteria are designed to actively engage in gene exchange by several complex and elaborate mechanisms. One of them is transduction, whereby temperate phages inject their DNA into bacteria.28 Laboratory research has found transduction can spread bacterial genes far beyond the locale where the virus incorporated the genes. Miller noted that:
‘… when a bacterium carrying a new gene enters a habitat, bacteriophages infect that cell and create more bacteriophage particles. If any particles end up containing the new gene, that gene can be passed on to the indigenous bacterial population. This model is equally applicable to the transduction of chromosomal and plasmid DNA. We have isolated bacteria and bacteriophages from various lakes and have demonstrated that bacteria do share genetic information by transduction in those settings. Many microbiologists originally thought transduction would not be an important means of gene exchange in the environment, because it requires viruses and bacteria—both of which were thought to be present in low concentrations—to interact. But … bacteriophages [exist] in very high concentrations (often 100 billion virus particles per milliliter) in fresh and marine waters. These observations have caused a re-evaluation of the frequency of interactions, including transduction, that occur between bacteriophages and their hosts.’29
The incorporation of plasmids or prophages into the bacterial chromosomes is called transfection. The incorporation of only genes by a virus carrier or free DNA is called transformation. Transfection and transformation are not random, but tightly controlled processes. Specific surface receptors determine which genes or gene packets enter the cell.
Although several ways exist in which genes can be transferred from one bacterium to another, transfer
‘occurs very rarely by transfection. In nearly all prokaryotes, it occurs commonly through the intermediary of an infectious form of prophage, the temperate bacteriophage or phage.’30
Plasmids rarely become integrated into a bacterial chromosome, but instead are like ‘book-mobile’ genes, passed around and used where needed, otherwise discarded (Figure 6). Not all viruses serve this function; many may serve some totally unknown function in the natural world. The realisation that some kinds of viruses have a wider role in life led to a revolution in biology. Using the past research as a guide, it is likely that the extant bacteria and viruses whose function we do not understand likewise will be found to have important roles in the natural world.
A strong argument for the gene carrier role (and other functional roles for viruses) is that viruses are comparatively simple, while cells are extremely complex and have elaborate defence systems. It is logical that cells would have elaborate defences, since these are necessary to prevent genetic take-overs. This is a major problem with evolution theory. In the words of Syvanen,
‘If viruses were nothing but bad news, you would expect cells to be taking great pains to evolve resistance to them and they do express resistance in some way but they also seem awfully accommodating in others.’31
Pathogenic viruses
The traditional understanding that viruses are alien invaders competing against humans in a life or death struggle for the cell’s manufacturing facility is now understood to be oversimplified, if not incorrect. It is usually not expedient for a virus to kill its host, since this may cause the death of the virus. Viruses must have a reservoir of host species in which they can live permanently otherwise they would soon go extinct. AIDS, for example, infects some primates without causing illness or death, and has probably lived in them in a commensal relationship for generations. The host organism must tolerate them fairly well—in fact, some kinds of viruses form a symbiotic relationship with their hosts.
Evidently, inappropriate sexual acts by humans caused the transfer of a lentivirus from a monkey to humans. As long as the HIV lentivirus lived in monkeys, it was not a threat for humans. HIV in monkeys (called SIV), ‘appears not to cause disease in most of its natural hosts’, and ‘bacteria and viruses that cause disease today may not always have done so’.32 The same situation also is true of syphilis (apparently from sheep) and many other infectious diseases. Baboons resist being adversely infected by HIV, and for years researchers have been exposing certain animals to the virus without infecting them.
This supports the argument that viruses normally do not, and should not, cause disease. Only if something goes wrong, such as a mutation or accidental inappropriate movement of genes, do they cause problems. Dr Charles Stiles recognised this many years ago when he concluded that ‘germs were not created as they are today, but they later evolved into germs … those germs were originally created in some form other than as disease germs.’33 Stiles claimed that germs developed as a result of the devolution that has occurred since creation.
Evidence is accumulating to suggest that most or all harmful viruses and bacteria are mutated forms of non-pathogens. Pathogenic organisms result from gene shuffling, which inadvertently disrupts the normal species-host relationship. Diphtheria is likely not the only case of a harmless organism becoming ‘pathogenic because of virulence genes brought by a prophage or plasmid.’34 The case of Vibrio cholerae (the causative agent responsible for the deadly disease known as cholera) illustrates this.
‘Just one strain of the bacterial species Vibrio cholerae wreaked almost all the death and misery. This strain is known as O1, and it produces a toxin that binds to cells of the small intestine, setting off a cascade of reactions in which cells pump out vast amounts of chloride ions and water—some five gallons a day. If salts and water are not quickly replaced the patient dies. Surprisingly, most strains of V. cholerae are harmless organisms that live and multiply in rivers and the open sea. But at some time in its evolutionary history, the O1 strain turned lethal. What caused this deadly transformation? A virus, according to microbiologist Mathew Waldor … [Waldor] and his colleague John Mekalanos at Harvard discovered the virus while studying the stretch of bacterial DNA known to include the gene called CTX, that codes for the cholera toxin. They suspected that a virus might have infected the bacteria with the gene, since viruses often insert their own genetic material into bacteria.’35
Later researchers confirmed that the cause was a virus. Although bacteriophages are similar to all other viruses, some differences exist. Viruses that function only as a parasite are defined by some researchers as true viruses, while bacteriophages are defined as non-viruses as some do not kill the cell but splice their DNA into the bacterial chromosome (see Figure 5). This terminology may have been coined to avoid some of the negative connotations of the word ‘virus.’ There is also some evidence that infectious agents still labelled viruses ‘carry favourable genes and help even in eukaryotic evolution.’ The research necessary to elucidate the origin of pathogenesis is complex, and the story
‘of how the once-harmless cholera bacterium got its toxin and so became a killer is … the all-too-real finding of years of sophisticated molecular detective work and its implications, … The discovery of CTX is just one of a string of surprises to emerge in the past few months from studies of the ways microorganisms interact with each other and their hosts to cause disease. From BSE [Bovine Spongiform Encephalopathy] to AIDS and malaria, researchers are confronting tough questions about the evolution of disease.’36
When bacteria were first discovered, few dreamed that they played such a critical positive role, as we now know they do in ecology—and that the same is also evidently true of viruses.37 Furthermore, viruses are everywhere in abundance. A major science story of the last decade relates to the discovery by marine biologists that a major proportion of the sea’s biomass is microscopic and is comprised of viruses, bacteria, algae, and protozoa.
‘A teaspoonful of seawater may contain more than a billion viruses—10,000 to 10 million times more than previously estimated. The world ocean is what laboratory scientists would call a culture medium, the largest petri dish known to mankind. There can be millions of individuals of a single species in an ounce of seawater, and presumably they play an enormous role in the planet’s carbon cycle. Whether Earth undergoes the global warming the world is watching for may be decided by organisms we didn’t know were there. As Bob Guillard of the Bigelow Laboratory of Ocean Sciences in Marine has observed: “A hundred years of oceanography, and the most abundant being in the world wasn’t recognized by anybody”.’38
Humans normally live in an environment inhabited by trillions of microorganisms and as many as 300,000 microorganism species. Estimates run as high as 10,000 for different species of bacteria and viruses per gram of soil.39 Many sea and land animals also live in a world of trillions of viruses, yet they rarely develop infections.40,41 It is estimated that of the cells that make up or live on a human body, fully 90% of them are microorganisms.42 Microorganisms are everywhere on, inside, and around us.
A body of evidence indicates that the presence of viruses is not, in itself, the primary cause of disease. This evidence comes from research on animals that live in environments that contain a high number of viruses and bacteria. One example that has been studied extensively is the shark. Mestel states: ‘You’ll rarely see a sick shark in the wild, although the oceans are teaming with bacteria and viruses …’.43 This is true in spite of the fact that sharks have simple immune systems that lack ‘T-cells’ (a type of immune system cell present in humans and animals), and thus only sluggishly reject tissue grafts.
Sharks also lack a complex antibody response such as that exhibited in humans. If foreign protein is injected into a shark, it will produce antibodies that bind the foreign antigens but, in contrast to humans, the immune response is not improved by repeated injections. Thus, sharks do not develop memory immunity.
It should not surprise us that in a post–Fall world, disease is a result of ‘something gone wrong’, as indicated from several lines of research, specifically research on
‘a range of “emerging” diseases that appear … to have entered the human population only recently. And here, perhaps more than anywhere else in the field, myths abound. The standard theories hold that, when a microorganism moves from one species to another—as HIV is believed to have done—it will be nastier than it had been in its original host. Again, this is simply wrong. According to Ebert, parasites tend to be less infectious, less fit and less harmful in new hosts. Of course, there are exceptions, and these are the ones we notice, says Ebert. In reality, bacteria, viruses and other parasites probably jump species far more often than anyone knows, without doing much damage.’44
Morse also states:
‘Common wisdom held that an emerging virus sprang forth so suddenly because it had evolved de novo … As it happens, the great majority of “new” viruses are not really new at all but are by-products of … viral traffic: the transfer to humans of diseases that exist within some animal population.’45
He also adds that ‘new’ viruses such as AIDS may have reflected only our imperfect knowledge of the natural world, not a radical new trend in viral evolution.46 The problem, he argues, is a biological mismatch—an organism transferred from its host to where it does not belong. In Morse’s words, by humans ‘disrupting the established ecological order, people inadvertently encouraged the adaptation of a “weed” species that more often than not brought them fever and misery.’47
This new understanding of pathogenesis origins is a major topic of scientific research, and is of special interest to creationists because this new evidence fits a creationist worldview. No doubt the Fall has contributed to problems developing in what once was a symbiotic, functional relationship between viruses and their hosts. Viruses may even be critical for the survival of certain life forms such as bacteria.
Pathogenic viruses are only the ‘tip of the iceberg’ of virus types, and the more that is learned about the biological world the more scientists are coming to realise the critical role that viruses play in life.48 The fact that so few kinds of microorganisms are pathogenic is evidence for the mutation theory of the origins of harmful viruses. Another piece of evidence is the fact that usually most strains are not pathogenic, with only one strain or a few strains causing problems. Furthermore, often viruses do not kill directly, but indirectly. For example, a hantavirus triggers a powerful immune attack by the host that may damage the host’s healthy cells.
Most pathogenic viruses are often more of a bother than a threat. Of the tailed bacteriophages, only 1% are virulent, and the only known viruses that are fatal in virtually every case where infection is established (in unprotected persons) are rabies and AIDS.49 The origin of this small fraction of pathogenic microorganisms is now a major focus of research.
The use of viruses in medicine
Bacteriophages (literally eaters of bacteria) may help to control bacterial growth and spread. Almost all known bacteria have a specific predatory phage. Research indicates that in certain animals some virulent viruses may fight against bacterial infections. The advantages of using viruses to treat illness, and the reason for the importance of such a concept was reviewed by Radetsky, who noted that few persons in the past
‘… wanted to fool around with live infectious viruses when you could pop a few penicillin pills … Western scientists bundled bacteriophage therapy into the dusty closets of history. Today it may be coming back. Some 50 years after antibiotics heralded the end of bacterial disease their golden age is waning … More and more microbes are developing resistance to our arsenal of antibiotic drugs, and scientists are again searching for miracle treatments. Some are looking to the past, to the almost forgotten bacteria eaters. In fact, bacteriophage therapy has never really disappeared. [Some] … doctors and health care workers routinely use bacteriophage therapy to cure a wide variety of maladies …’50
The clear advantages of virus treatment include:
‘… even if antibiotic–resistant bacteria were not such a burgeoning threat, phage therapy would still be appealing. Antibiotics involve certain risks. They kill a wide range of bugs, not just their particular targets, and so rid the body not just of harmful microbes but of useful ones as well—bacteria that aid in digestion, for example. For antibiotic therapy to be effective, patients must diligently take multiple doses over an extended period of time. Slack off and you may find yourself battling a renewed attack of disease, this time borne by resistant bacteria. Antibiotics can cause intestinal disorders and yeast infections. Finally, some people are violently allergic to antibiotics. In such cases, the cure can be worse than the disease. None of these problems apply to bacteriophages. Phages prompt no allergic reactions and are notoriously finicky—they target only the bugs they’re supposed to. And if you miss a dose of phage, no problem. Because they reproduce within the bacteria they attack, they stay around for a few days before the body can clear them from the system.’51
Though not without problems, the technique holds much promise. Some examples of alleged success include treatment of dysentery, typhoid fever, food/blood poisoning, and skin/throat/urinary-tract infections:
‘If someone has an intestinal disorder, the person can drink the phage … If it is a skin infection, phage can be applied to the spot. We have developed aerosol and tablet preparations.’52
Bacteriophages usually attach onto the surface of only one specific bacterial species, a fact that can be used to distinguish between bacterial strains (a process known as bacterial typing).
One potential example of the use of viruses to cure disease is to employ a virus to kill a virus. A virus can become a biological weapon to seek out HIV infected cells. A benign virus coated with special proteins can seek out cells infected with HIV, and then lock onto the cell’s surface. The HIV virus carries molecules that link with receptors on the surface of cells that it infects. When the HIV molecule connects with the receptor, it allows the virus to enter. A primary receptor used by HIV is CD4, the receptor present on immune system blood cells that are the primary targets of HIV. HIV also requires the use of at least one or two other receptors present on immune system cells. One is a receptor called CCR5, which is used by HIV early in the disease to infect a macrophage. Another receptor, the CXCR4, is used by HIV later to infect T-cells.
Researchers used this information to coat the surface of a harmless virus with the molecules used by HIV to invade cells. The altered virus then was exposed in the laboratory to HIV-infected cells. The hunter virus coated with CD4 and CCR5 successfully locked onto HIV-infected macrophages. When coated with CD4 and CXCR4, the hunter virus sought out and locked onto HIV infected T-cells. In both cases, the hunter virus ignored normal cells that were not infected with HIV. ‘This approach could offer a way to deliver antiviral genes directly to HIV-infected cells in vivo …’53
Viruses and the genetic revolution
An infected animal cell can express thousands of copies of many kinds of proteins, but can produce only enough viral proteins for as many as six viruses. On the other hand, if the viral protein genes are spliced into a bacterium’s DNA by recombinant DNA means, the bacterium will mainly manufacture these viral proteins, making it far easier to separate and study them. This greatly simplifies the process of genetic research. Another advantage of using viruses in research is that virtually all viruses of a particular strain are identical.
Just how critical viruses are for medical and molecular research is now obvious to all molecular biology researchers. Zimmerman and Zimmerman (1993) noted that in molecular biology ‘today is the day of the virus’ and ‘nothing being studied in medicine, nothing in biology, is more important.’54 Among the many tools that are critical in molecular biology, and that were either discovered in viruses or exist because of viruses, include: reverse transcriptase, restriction enzymes made by bacteria to control viruses, and many other enzymes. This research has aided virologists enormously in exploring the relationship between the virus and its host, and the mechanism of pathogenicity.
The gene carrier function of viruses might soon be brought into service to perform a critical role in curing genetic diseases. Genetic defects cause over 5,000 known diseases, including Huntington’s chorea, sickle cell anaemia, and cystic fibrosis. The present goal of gene therapy, which was tried first with cystic fibrosis, is to load virus-carriers with healthy genes and then infect the relevant tissues so that the cell incorporates the new virus-carried genes into its own DNA. Viruses are ‘the ideal way to transport genes because they naturally infect cells to deposit genetic material.’55
The most common vector virus, Moloney Murine Leukaemia Virus, has been used in about three-quarters of gene therapy treatments tried so far. Viruses modified by adding the genes for the enzyme cre–recombinase effectively cause the virus to ‘commit suicide after dispatching the therapeutic gene.’56 Specifically, the cre–recombinase cuts out the viral DNA, leaving behind only the therapeutic genes. This reduces the likelihood that the virus will cause problems, or that its genes will be spliced in the wrong place.
Viruses are still a source of major breakthroughs in genetic therapy. Researchers now have found a virus that
‘… likes to hang around on its own … [which] has solved a vexing problem for scientists who have to shuttle foreign genes into mice … The technique … could vastly accelerate and simplify the analysis of new genes. Genetic research took a great leap forward when biologists discovered more than a decade ago that they could incorporate new DNA into a developing mouse embryo simply by injecting it straight in. But this technique suffers from one big drawback. Many millions of copies of a gene are injected into the embryo at once, and for unknown reasons they tend to mingle and form a long chain of genes before splicing themselves into the mouse chromosome.’57
This virus may solve this aggregation problem because it usually infiltrates the chromosomes one at a time, evidently due to proteins that bind at the end of DNA and function as caps to prevent the viral copies from linking together.
Virus therapy is especially promising for some neurological disorders (including Alzheimer’s and Parkinson’s), and many inherited disorders and types of brain tumours. A major difficulty in treating brain disorders is that many substances simply will not cross the blood/brain barrier. Certain viruses can pass through the blood/brain barrier to carry new genes to restore health. More than 100 different clinical trials are under way to research this very promising therapy.58 Lentiviral vectors seem especially promising. Modified Herpes virus strains have already been used to treat brain tumours successfully in mice. This particular research used a virus that multiplied inside the tumour cells, producing an enzyme to function as a catalyst to convert a non-toxic drug into a compound that destroys only tumour cells.59
The immune system
The immune system of vertebrates usually is a highly effective defence against nearly all pathogenic viruses. When things go wrong, the problem often is caused by factors such as an immune system weakened by a genetic mutation, poor diet, poor sanitation, exposure to new pathogens (or such high numbers of pathogenes that the immune system is overloaded), emotional stress, lack of sleep or lack of exercise. Some viruses have the ability to change their antigenic structure so that the body’s defence system perceives it as a new, unknown foreign organism.
Viruses generally do not evolve or mutate to overcome their hosts’ immune systems, but as Morse notes, the crucial event in generating new strains ‘has not been mutation but the successful reshuffling of preexisting genes.’60 Many of these genes may have been transferred from animals. Thus their pathogenesis may be due not to design, but rather to an accident. Consequently, the host must develop a completely new immune response to the invaders—which takes time. For this reason, the acute phase of most illnesses lasts at least 2–3 days—until the body’s defence response can develop sufficiently to destroy the invaders. Flu, colds (rhinovirus infection), and the AIDS virus are more adept than most viruses at using this genetic process of reshuffling genes to produce new strains.
This reshuffling, often referred to incorrectly as a mutation, usually is a designed response to allow survival of the virus. Under ordinary circumstances, certain genes manufacture surface proteins that enable the host’s immune system to identify a virus as foreign. However, when shuffling of genes by a virus results in the production of new antigenic proteins, the virus is not immediately recognised by the immune system, thus slowing down the immune response. This gene reshuffling also occasionally may splice genes in the wrong place in the genome, producing pathogenic bacteria or viruses as discussed above.
Beyond these programmed changes, we have no clear evidence of the evolution of viruses. All the extant evidence indicates that ancient viruses are identical to those found today.61
The body’s defence system involves more than just the immune system. The skin excretes RNase, which cuts up the RNA in RNA-based viruses. The skin also secretes magainins, which can kill pathogens. This complex system almost always is effective; thus, the trillions of viruses all around us rarely cause problems.
Summary
Virology is a relatively new field of study. Many researchers have concluded that we are now at the same point in our understanding of viruses that scientists were in researching bacteria at the turn of the 20th century. Viruses now are known to serve several beneficial roles, and research has indicated several others may be important. According to this model, disease is not a result of viruses as much as a breakdown of a virus/host relationship. Thousands of virus types exist in host cells without problems. Problems that do cause disease are a result of reshuffling of virus genes, genetic mutation of the host, or a breakdown in the general health of the host organism. Research is showing that viruses are a critical part of life. Holmes has noted:
‘For sheer numbers, no other ocean beings can match viruses. Thousands, sometimes even millions of these molecular parasites inhabit every drop of surface seawater, outnumbering even bacteria by 10 to 1 … evidence that suggests that viruses are a powerful force in the sea, and one that determines how many plankton and ultimately how many fish, and even humans, an ocean ecosystem can support … viruses must have a profound influence on the entire oceanic ecosystem. When protozoans eat bacteria, energy passes along the food chain leading from protozoa to other zooplankton to larger predators, including fish. But when virus-infected bacterial cells burst, their energy-rich cell contents spill into the water for other bacteria to scavenge. “Viruses tend to keep nutrients away from the big stuff and keep them going around in the little stuff,” says Fuhrman. If so, viruses have shaped the entire structure of the ecosystem.’62
Acknowledgements
I wish to thank both Bert Thompson and John Woodmorappe for their suggestions and extremely helpful feedback.
References and notes
- Woodmorappe, J., Noah’s Ark: A Feasibility Study, ICR, Santee, CA, 1996. Return to text.
- Young, W., Fallacies of Creationism, Detselig Enterprises, Calgary, Alberta, 1985. Return to text.
- Zimmerman, B. and Zimmerman, D., Why Nothing Can Travel Faster than Light, Contemporary Books, Inc., Chicago, IL, 1993. Return to text.
- Hsiung, G.D., Diagnostic Virology, Third Edition, Yale Univ. Press, New Haven, CT, 1982. Return to text.
- Curtis, H., The Viruses, The Natural History Press, Garden City, NY, 1966. Return to text.
- Stanley, B., Animal Viruses, Vol. 3 Academic Press, Inc., New York, 1959. Return to text.
- Valentine, R.C. and Pereira, H.G., Antigens and structure of the adenovirus, J. Mol. Biol. 13(43):13–20, 1965. Return to text.
- Jensen, M., Wright, D. and Robinson, R., Microbiology for the Health Sciences, Prentice Hall, Upper Saddle River, NJ, 1996. Return to text.
- Gallo, R.C., The AIDS Virus, Scientific American 256(1):37–48, 1987. Return to text.
- Starr, C., Biology; Concepts and Applications, Wadsworth Pub. Co., Belmont, CA, p. 292, 1996. Return to text.
- Simon, L.D. and Anderson, T.F., The infection of Escherichia coli by T2 and T4 bacteriophages as seen in the electron microscope, I. II. Virology 32(158):279–305, 1967. Return to text.
- Coombs, D. and Ariska, F., T-4 Tail Structure and Function, p. 28, 1994. Return to text.
- Brooks, S., The World of Viruses, A.S. Barnes, London, 1973. Return to text.
- Talaro, K. and Talaro, T., Foundations in Microbiology, Wm C. Brown, Dubuque, IA, p. 132, 1993. Return to text.
- Evans, A.S., Viral Infections of Humans, Third Edition, Plenum Publishing Corp, New York, 1989. Return to text.
- Stent, G.S., and Claendar, R., Molecular Genetics: an Introductory Narrative, W.H. Freeman and Company, San Francisco, p. 319, 1978. Return to text.
- Starr, C., ref. 8, p. 294. Return to text.
- Hapgood, G., Viruses Emerge as a New Key for Unlocking Life’s Mysteries, Smithsonian 18(8):126, 1987. Return to text.
- Margulis, L., Foreword to Sonea and Panisset, p. vii, 1983. Return to text.
- Margulis, ref. 16. Return to text.
- Karam, J.D. (editor), Molecular Biology of Bacteriophage T-4 ASM Press, Washington, D.C., 1994. Return to text.
- Sonea, S. and Panisset, M., A New Bacteriology, Jones and Bartlett, Boston, MA, 1983. Return to text.
- Mathieu, L. and Sonea, S., A Powerful Bacteria World, Endeavor 13(3):115, 1995. Return to text.
- Miller, R., Molecular Biology of Bacteriophage T-4, ASM Press, Washington D.C., 1998. Return to text.
- Mathieu, ref. 20, p. 112. Return to text.
- Hecht, J., Rare Bug Dominates the Oceans, New Scientist 144(1952):21, 1994. Return to text.
- Sonea and Panisett, ref. 19, p. 36. Return to text.
- Mathieu and Sonea, ref. 20, p. 114. Return to text.
- Miller, ref. 21, p. 71. Return to text.
- Sonea and Panisett, ref. 19, p. 42. Return to text.
- Hapgood, ref. 15, p. 126. Return to text.
- Brown, P., How the parasite learnt to Kill, New Scientist 152(2056):32–36, 1996. Return to text.
- Quoted in The San Antonio Express 1 April 1923. Return to text.
- Mathieu and Sonea, ref. 20, p. 112. Return to text.
- Glausiusz, J., How Cholera Became a Killer, Discovery 17(10):28, 1996. Return to text.
- Brown, ref. 29, p. 32. Return to text.
- Mathieu and Sonea, ref. 20. Return to text.
- Wiley, J., Phenomena, Comment and Notes, Smithsonian 21(4):29, 1990. Return to text.
- Tiedje, J.M., Microbial Diversity: Of Value to Whom? American Society for Microbiology News 60(10):524, 1994. Return to text.
- Homes, B., Who Rules the Waves? New Scientist 152(2054):2 supp, 1996. Return to text.
- Mestel, R., Sharks’ Healing Powers, Natural History 105(9):40–48, 1996. Return to text.
- Creager, J., Black, J. and Davison, V., Microbiology Principles and Applications, Prentice Hall, Englewood Cliffs, NJ, p. 2, 1990. Return to text..
- Mestel, ref. 38, p. 41. Return to text.
- Brown, ref. 29, pp. 35–36. Return to text.
- Creager, ref. 39, p. 16. Return to text.
- Creager, ref. 39, pp. 16–18. Return to text.
- Creager, ref. 39, p. 18. Return to text.
- Zimmerman and Zimmerman, ref. 3. Return to text.
- Curtis, ref. 5, p. 14. Return to text.
- Radetsky, P. The Good Virus, Discover 17(11):52, 1996. Return to text.
- Radetsky, ref. 47, pp. 54–55. Return to text.
- Radetsky, ref. 47, p. 56. Return to text.
- Enders, M., et al., Targeting of HIV-and SIV-Infected Cells by CD4-Chemokine Receptor Pseudotypes, Science 278:1462–1463, 1997. Return to text.
- Zimmerman and Zimmerman, ref.3, p. 274. Return to text.
- Hotz, R., Researchers Alter Viruses to Combat Brain Disorders, The Journal Gazette, pp.2, 20 November 1996. Return to text.
- Coghlan, A., This Message Will Self-Destruct … , New Scientist 151(2042):20, 1996. Return to text.
- Cohen, P., Lone Virus Speeds the Gene Shuttle, New Scientist 151(2044):23, 1996. Return to text.
- Hotz, ref. 52, p. 1. Return to text.
- Hotz, ref. 52, p. 2. Return to text.
- Morse, S., Stirring up Trouble; Environmental Disruption Can Divert Animal Viruses into People, Science 30(5):20, 1990. Return to text.
- For a discussion of this, see Herrmann, B. and Hummel, S., Ancient DNA, Springer, New York, NY, 1994. Return to text.
- Holmes, B., Who Rules the Waves? New Scientist 152(2054):8–9, supp, 1996. Return to text.
- Luria, S.E., Darnell Jr., J. E., Baltimore, D. and Campbell, A., General Virology, John Wiley and Sons, New York, NY. p. 36, 1978. Return to text.
- Luria, ref. 63, p. 159. Return to text.
- Luria, ref. 63, p. 228. Return to text.






Readers’ comments
Comments are automatically closed 14 days after publication.