

Journal of Creation 32(3):88–96, December 2018
Browse our latest digital issue Subscribe
Paleoanthropology in Australia—Homo erectus and modern human origins
Robust human fossil remains have been found in Australia, at sites like Kow Swamp and the Willandra Lakes region. Their identity in relation to specimens of Homo erectus, and their role in human ancestry, has been a controversial one. This issue will be addressed in this article, as well as models of modern human origin that attempt to explain the robusticity seen in early Australians. Also touched on is the politics that has seemingly paralyzed much of Australian paleoanthropology for decades. The Out of Africa and Multiregional evolutionary theories of modern human origins are considered, as well as an explanation based on biblical events.
Homo erectus

Eugene Dubois, responsible for the 1891 discovery of the first Homo erectus specimen (known back then as Pithecanthropus erectus), in Java, Indonesia, claimed that his “Pithecanthropus was a ‘venerable ape-man’”.1 A more current view of Homo erectus, by evolutionists, is that it is “an early-to-mid-Pleistocene species thought to be on the direct ancestral line to modern humans”.2 According to Campbell and Loy, Homo erectus individuals “seem to have a modern body build”, in regard to the postcranial skeleton, but “differed greatly from modern people in their brain size and cranial anatomy”.3 Paleoanthropologist Daniel Lieberman writes that “In essence, H. erectus was the first ancestor we can characterize as significantly human.”4 Homo erectus expert Susan Antón states:
“… our opinion of H. erectus has changed as our understanding of human evolution has matured. Initially seen as perhaps no more than a fossil ape or a regional isolate, H. erectus is now considered the hominin that was the first to take major anatomical and behavioral steps in the direction of a ‘modern human’ body plan.”5
According to paleoanthropologist Chris Stringer:
“Most anthropologists recognize the existence of at least two human species during the last million years—the extinct Homo erectus and our own species, Homo sapiens—but there are very different views on how these species are related.”6
Whilst evolutionists may refer to Homo erectus as a human species, what that means is generally different from the belief of most creationists, that Homo erectus individuals were fully human and hence descendants of Adam and Eve. This paper is not so much about the features of Homo erectus, but rather, considers evidence for erectus-like fossil specimens having been found in Australia. While finding Homo erectus fossils in Australia would not be a problem for the creationist position, it does cause problems for evolutionary theories of modern human origins that regard Homo erectus as a less evolved human. One reason for this is that, according to Bae et al.:
“It has long been argued that modern humans were the only hominin taxon capable of peopling Australasia, particularly because it would have involved the ability to build sturdy watercraft and navigate the open seas.”7
Homo erectus showing up in Australia would be a major problem for evolutionary models of human origins, such as the Out of Africa (OoA) theory, because, as indicated above, this model suggests that Homo erectus did not have the intelligence to make well-built watercraft and navigate the open seas, which would be required to reach Australia. It also makes it harder to argue that Homo erectus individuals were less intelligent than modern humans.
An even bigger problem, though, is that if the robustly built humans known to have been in Australia were erectus- like, and if Australian Aborigines, as believed, are linked to (i.e. descended from) both the gracile (anatomically modern) and robust (i.e. erectus-like) people, as well as people with a “more middle of the road morphology”,8 what does that mean if Homo erectus is not considered fully human (i.e. a member of Homo sapiens)? Regarding questions like these, evolutionists from the multiregional school of thought have expressed concerns about OoA (or Eve) theorists defining the species Homo sapiens too restrictively, as follows:

“We believe that an unfortunate aspect of the debate are definitions of Homo sapiens used by some ‘Eve’ theorists. They have been found to exclude many Pleistocene and more recent Aboriginal Australians from our species … . Further examination of these individuals and collections of recent Aboriginal skeletal remains leads us to estimate that these definitions of modern Homo sapiens exclude anywhere between 40 000 and 60 000 living Aboriginal Australians. We feel that there is great danger in this. It is the duty of specialists to make sure that they include all living people in any definition of our species. If we do define humans as minimally including all living people, many of the fossils that the ‘Eve’ theorists claim leave no descendants, including Neanderthals, fall into Homo sapiens.”9
Kow Swamp
When Thorne and Macumber described the human remains from Kow Swamp, Australia (the first remains were discovered in 1967; figure 1) in the journal Nature, in 1972, they wrote:
“Analysis of the cranial morphology of more than thirty individuals reveals the survival of Homo erectus features in Australia until as recently as 10,000 years ago.”10
From this it might seem that the question of erectus- like fossils existing in Australia was already answered in the affirmative back then, but a changing political climate in the 1980s, as well as the increasing influence of the OoA theory of modern human origins during the same decade, meant any such claims would be challenged. For example, it has been suggested that “the ‘robust’ Australian crania that look the most archaic, like Cohuna and Kow Swamp 1 and 5, have been shown to share strong shape similarities with known artificially deformed crania”.11 Commenting on suggestions that some specimens from Kow Swamp (KS) had been artificially deformed, Darren Curnoe stated:
“However, a recent study (Curnoe and Thorne, 2006b) [see Curnoe and Thorne12] has found no evidence for such modification in KS 1 [figure 2] and the case for artificial deformation in KS 5 [figure 3] to be at best very weak. Other relevant specimens from Australia such as Cohuna, WLH 50, KS 8, KS 9, Mossgiel, LM 3 and King Island are incomplete and many standard measurements cannot be reliably taken on them (Curnoe and Thorne, 2006a) [see Curnoe and Thorne13].”

Other researchers, including Arthur Durband, disagree with Curnoe’s assessment, with Durband stating that the results from his analysis “show that Kow Swamp 1 and 5 have strong shape similarities to known deformed individuals”.14 Reporting on a “new discriminant function for the identification of artificially deformed crania”, Clark et al. scored Kow Swamp 5 as artificially deformed, but not Kow Swamp 1.15 From their results the authors stated:
“These results provide further support for the argument that deformation is present in some specimens in the early Australian sample, and, as such, it is not appropriate to use cranial characteristics ‘shared’ between Indonesian hominids and early Australians with evidence of deformation to infer linkages between the two populations.”16
While one can agree with the above statement, it should not mean that crania that do not appear to be artificially deformed (e.g. Kow Swamp 1) cannot be used as evidence of linkage between populations, if indeed that is what it shows. There is disagreement about whether (or which) Kow Swamp specimens show possible evidence of having been artificially deformed, but getting a definitive resolution to the argument may be out of reach. This is because of the policy of repatriation of ancient remains that was starting to take place in the 1980s which, concerning the Kow Swamp fossil remains, and according to John Mulvaney and Johan Kamminga, led to the Museum of Victoria in 1990 unconditionally returning “the entire collection to the Echuca Aboriginal community and presently its fate remains obscure”.17
Changing political climate
Regarding the changing political climate alluded to, Arthur Durband and Michael Westaway have described the situation as follows:
“Unfortunately, work on the prehistory of Australia was significantly impacted in the 1980s by changes in legislation surrounding the ownership of ancient remains … . Control was transferred to local Aboriginal groups, who reclaimed and reburied many of the Pleistocene human remains recovered by that point … . It also became much more difficult to publish data or photographs of ancient Australians, as control over the use of any data or images of the fossils also became subject to the discretion of the local Aboriginal groups. As a result, paleoanthropological fieldwork in Australia essentially ground to a halt as much of the modern debate over the origins of modern humans was beginning to take shape.”18
Independent researcher Vesna Tenodi has written a less diplomatic view of the situation, as follows:
“Ancient Australian skulls can not be investigated, nor reconstructed. Replicas or even drawings cannot be displayed, or discussed, as that also is too offensive and cannot be done without ‘Aboriginal permission.’ Even when skulls are clearly non-aboriginal, such as the Mungo Man or the Kow Swamp skeletons.
“My conclusion is that all these enforced ‘protocols’ were invented to protect the political decision to maintain the dogma that Aborigines are the ‘First people’.”19
Given the above-mentioned situation with respect to Australian human fossils, one essentially has to rely on descriptions of fossils discovered before the change in political climate, such as the Kow Swamp and Willandra Lakes region fossils, to answer the question of whether erectus-like individuals were once in Australia. In discussing Australian skulls possibly affiliated with Homo erectus, including the Talgai skull, Pintupi skull, and the Kow Swamp and Coobool Creek skulls, Tenodi, who also describes herself as a ‘spiritual archaeologist’, believes the evidence “points at the parallel existence of both Homo erectus and Homo sapiens, and indicates the presence of much older, highly-evolved pre-Aboriginal cultures”.20
Single Origin Model

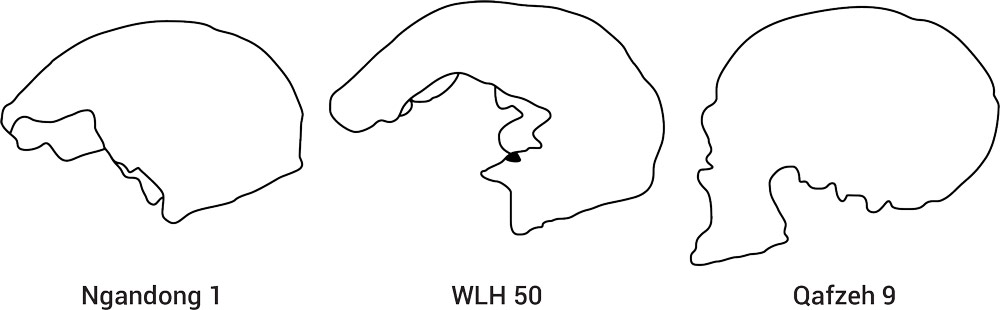
Westaway writes that there are “very lightly built” (gracile) crania in the Willandra Lakes region, such as Mungo Lady (also known as Lake Mungo 1 or WLH 1) and Mungo Man (Lake Mungo 3 or WLH 3; figure 4), and there are also fossils in the same region from “very heavily built individuals”.21 The term ‘robust’ is often used in describing these specimens and the fossil humans from Kow Swamp, Victoria.21 Their thick cranial vaults, large brow ridges and “pronounced areas of muscle attachment” are said to be “cranial distinguishing markings” of these robust individuals, the best known being Willandra Lakes Human 50 (WLH 50). 21 Hence, any Australian human origin model has to explain both the robust and gracile human forms.
The evolution-based dating methods give inconsistent dates for some of the Australian fossil remains, depending on the method used. The gracile specimen Lake Mungo 3 is dated as either 40 ka (on geological grounds) or about 61 ka (ESR and U-S dating), both of which are older than the Kow Swamp dates of about 13 to 9.5 ka (radiocarbon dates on shells) or 26 to 19 ka (optically stimulated luminescence).22 Therefore, any model that accepts the ages given by these dating methods needs to also explain why a gracile human at the Willandra Lakes region appears to be dated earlier than the robust humans at Kow Swamp. The most mainstream evolutionist position to explain the gracile and robust groups of early Australians is probably the Single Origin Model (SOM), which proposes “that a single founding group initially populated Australia”, as described by Durband and Westaway:
“Instead, the observed range of biological variation can be accounted for by localized evolutionary processes combined with cultural factors like mate selection that would increase the potential for genetic drift. Additionally, cultural phenomena such as interpersonal violence and artificial cranial deformation also influenced cranial shape and thickness, accentuating some regional distinctions between populations. In short, a single origin model for the Australians, combined with the effects of selection, is perfectly consistent with the evidence and presents a parsimonious scenario.”23
Proponents of the SOM are likely to be supporters of the Out of Africa (OoA) theory of modern human origins, who are said to “argue that the variation within the fossil sequence in the Willandra region represents the extremes of a homogenous population across Australia”.24 For the SOM to work, skulls similar to Homo erectus, like WLH 50, described by Durband and Westaway as “an exceptionally massive skull, one of the most robust modern humans yet discovered”, must be explained away or downplayed, and so they state:
“While it is often highlighted for its unusual, extreme morphology, it is because of this singular anatomy that WLH 50 is arguably of less importance for our understanding of modern human origins than any other single specimen in Australia.”25
It is circular logic, and convenient, to say that the fossil which arguably is the most difficult for the single model to explain, is the least important because of its ‘extreme morphology’. Peter Hiscock stated, “It seems reckless, as Brown … and Cameron and Groves … have argued, to base any interpretation of human evolution on this unusual, pathological individual”, claiming it was likely that WLH 50 “suffered an illness that changed his crania [sic]”, yet admitted “his particular problem has not been diagnosed”.26 Needless to say, the advice of ignoring WLH 50 will not likely be heeded by some researchers, and will not be adhered to in this paper. Durband and Westaway also write:
“As WLH 50 is clearly unusual in its morphology, falls outside the normal range of variation for Pleistocene Australians, is likely pathological to some degree, and probably dates to at least 13 kyr after the initial colonization of Australia, it is perplexing that this individual has received this degree of notoriety in the debate over the origins of modern humans in Australia.”27
Dihybrid Model
One of the competing mainstream evolutionary positions, supported by proponents of the Multiregional Continuity Model (MCM) of modern human origins, is that Australia was populated by two groups of humans, “each with a distinct evolutionary origin, that coexisted for long periods of time in the late Pleistocene”.28 As stated by Glenn Conroy and Herman Pontzer:
“According to this theory, one group is represented by the robust Talgai, Cohuna and Kow Swamp specimens, which are seen as being derived from Indonesian H. erectus through such intermediaries as Ngandong. The second group is represented by the more gracile Mungo and Keilor fossils, which are seen as being derived from the Chinese H. erectus populations.”29
This dual migration model, where “two very different populations arrived and intermixed to create the diversity of Aboriginal people”, is known as the Dihybrid Model, and was advocated by the late Alan Thorne, who also linked it to the Multiregional Continuity Model of human evolution.30 According to Hiscock, the multiregional model:
“… hypothesized that modern humans evolved from earlier hominids in a number of regions, each with specific genetic traits but unified as a single, global species by inter-regional gene flow”.31
Willandra Lakes Human 50 (WLH 50)

The WLH 50 remains were discovered on the surface, not in situ, near Lake Garnpung, in the Willandra Lakes region (figure 5), in 1980.32 Regarding the WLH 50 cranium, there does not appear to be any claims about it having been artificially deformed, but as indicated earlier, it has been suggested that “its cranial vault profile is also likely anomalous due to pathology”.33 The age of WLH 50 is said to lie between 10.4 and 37.4 ka (thousand years ago), but most likely about 26 ka.34 According to the authors of the dating analyses:
“There is general agreement that WLH 50 is a modern human … . However, some see strong resemblances between WLH 50 and the Ngandong specimens … , whilst others attribute its robusticity to a pathological source … . However, detailed new studies refute the evidence for pathological hyperostosis of WLH 50 … . In a recent, comprehensive comparison of the metric and nonmetric features of WLH 50 with Levantine, African, and Ngandong specimens, Hawks et al. (2000) [see Hawks et al.35] concluded that WLH 50 was most closely related to the Ngandong specimens and that its robusticity was clearly attributable to its archaic rather than its pathological nature. Their conclusion, following Thorne and Wolpoff (1992) [see Thorne and Wolpoff36], was that WLH 50 belonged to the same species as Ngandong and was an example strongly endorsing the evolutionary model of regional continuity.”34
From their analyses, indicating anatomical similarities, Hawks et al. concluded that “WLH-50 and other Late Pleistocene fossils are modern humans, and the clear implication of their links to Ngandong is that these older Indonesians are H. sapiens as well.”37 Steve Webb, who originally suggested pathology may explain (in part), the “exceptional cranial vault thickness” of WLH 50, writes:
“It is clear that WLH 50 is a very robust individual with or without pathology, because other heavily developed features are clearly not pathological. Beside any pathology that might have moderately exaggerated its vault thickness, this individual would have had a thick vault in keeping with its other robust features.”38
Even if WLH 50 had some pathology (which seems doubtful), there are other Willandra Lakes individuals that have, for example, cranial vault thicknesses and well-developed (robust) brow ridges within the Homo erectus range.39 In the opinion of Webb, who has studied and described much of the fossil material recovered from the Willandra Lakes region:
“The distinct morphology of some Australian fossil crania cannot have an origin anywhere else other than Indonesia, it is not something developed within Australia, an aberration of the Ice Age, nor can it be one end of a wide range of anatomically modern human morphology that came ‘Out of Africa’. Indeed, it was precisely that anatomically modern humans were not cranially robust that distinguishes them as such. Whatever the genetic evidence suggests, the only people around to convey such an outstanding morphology must have been Homo soloensis or its descendants. There is no other Upper Pleistocene population from whom such a heavily built cranial structure could be inherited.”40
Note that Homo soloensis is used by some when referring to the Ngandong Homo erectus series crania. According to Josephine Flood:
“WLH 50 is massive: he is so robust, he makes Kow Swamp man look gracile! The cranium is extremely wide and approximately 210 millimetres long. The cranial vault bone averages 16 millimetres thick. Massive brow ridges form a continuous torus above the eyes, and the forehead is flat and receding. The back of the skull shows even more archaic characteristics, with substantial cranial buttressing. The neck muscle area is huge, the skull is extremely wide, the greatest width occurs very low in back view, and the difference between the width above and below the ears is much greater than in any modern people. Yet WLH 50’s brain was extremely large; the estimated endocranial volume is 1540 millilitres, well above the average 1300 for modern skulls.”41
Clearly WLH 50, “regarded as a modern human in all origins models”,42 is a very robust skull. It closely resembles the Indonesian Ngandong (Solo Man) Homo erectus series crania, such as Ngandong 1 (see figure 6). There should not be this resemblance, though, if Solo Man was derived from Homo erectus ‘hominids’ that left Africa nearly 2 Ma (million years ago). If the OoA theory is correct WLH 50 should most closely resemble ‘early’ anatomically modern human crania, such as Qafzeh 9, the type it is supposedly derived from, but it does not, as reported by Wolpoff et al.43
The Out of Africa (OoA) Model
From an evolutionary perspective the OoA model has been the most popular and influential theory of modern human origins in the last two or three decades. The OoA model proposes that modern humans emerged out of Africa about 100,000 years ago, with the Neanderthals replaced by the spread of these modern humans into their regions, and
“Like the Multiregional Model, this view accepts that fossils assigned to Homo erectus evolved into new forms of human in inhabited regions outside Africa, but argues that these non-African lines became extinct without evolving into modern humans.”44
The key points of the Out of Africa theory are summarized by Colin Barras:
“Most anthropologists believe that our species arose in Africa 200,000 years ago. Our genes show we are all descended from a single population that left Africa within the last 120,000 years and went global. This African group is the source of all human genes, barring a few gained by mating with species like Neanderthals.”45
While a pure OoA scheme allows no interbreeding between the emerging African modern humans and ‘resident archaic people’, interbreeding is allowed to different extents in variants of the OoA theory.46 The OoA model has fallen on hard times in recent years, particularly the pure replacement version of the model, as evidence of introgression between modern humans and supposedly more ‘archaic lineages’, like Neanderthals, have been piling up.47 According to Posth et al., the mitochondrial DNA (mtDNA) in a femur “displaying archaic hominin morphology”, from Germany, dated by them to 124 ka, indicated there had been “African mtDNA gene flow into Neanderthal populations”, the introgression event said to have occurred between 460 ka and 219 ka.48 Reporting on the study, Gibbon stated:
“After comparing the mitochondrial DNA (mtDNA) with that of other archaic and modern humans, the researchers reached a startling conclusion: A female member of the lineage that gave rise to Homo sapiens in Africa mated with a Neandertal male more than 220,000 years ago—much earlier than other known encounters between the two groups.”49
From an evolutionary point of view, the above 2017 study by Posth et al. seems to put early modern humans in Europe, which they constrain as “taking place more than ~270 ka”,50 at a time not only before they left Africa, but even before they had supposedly evolved in Africa (about 200 ka). This should be fatal to the OoA theory, no matter how much interbreeding is allowed in spin-offs of the theory, but, ironically, it is the fall of another key premise of the OoA tale that gives it some respite.
In 2017, a paper was published about a Homo sapiens fossil skull from Jebel Irhoud, Morocco, that was re-dated from about 160 ka to a much older 315 ka,51 and so, purely from an evolutionary point of view, humans may have appeared much earlier in Africa than thought, and it may not have been in East Africa that they first appeared. This raised doubts about other aspects of the OoA theory, as some evolutionists likened the Jebel Irhoud skull to the Dali skull (sometimes affiliated with Homo heidelbergensis or Homo erectus) from China, dated to 260 ka, with confusing implications:
“It’s possible, she says, that the hominins in Africa weren’t cut off from those in Eurasia. The small-scale movements of individuals—like young adults leaving a family group and joining a neighbouring one—could have allowed genes to flow across Africa and Eurasia. That means the genetic features of H. sapiens that appeared in Morocco 315,000 years ago could have cropped up in individuals—like the Dali skull’s owner—in China 260,000 years ago. There is another implication. ‘I think gene flow could have been multidirectional, so some of the traits seen in Europe or Africa could have originated in Asia,’ says Athreya [researcher at Texas A&M University]. That means features associated with H. sapiens may have evolved in east Asia, and been carried to Africa. If so, our origins are not solely African.”45
Even more recently, in 2018, a maxilla and associated dentition discovered at Misliya Cave, Israel, whose morphological traits were consistent with it being Homo sapiens, were dated from 177 to 194 ka, an age said to suggest “that members of the Homo sapiens clade left Africa earlier than previously thought”.52 Prior to this report, according to the authors, “the earliest modern human fossils found outside of Africa” were dated to around 90 to 120 ka, at the sites of Skhul and Qafzeh, also in Israel.53 The only thing that can be said with any degree of certainty is that the preferred Out of Africa theory of modern human origins is in a state of confusion and contradiction, as central tenets of the theory have toppled like dominoes, one by one.
The Multiregional Continuity (MC) Model
The evolutionary alternative to the Out of Africa Model, known as the Multiregional (or Multiregional Continuity) Model, argues “that ancient ancestors of various human groups lived where they are found today”.54 In this view humans, as in Homo erectus, migrated out of Africa nearly two million years ago, to different regions of the world. But rather than being replaced by subsequent recent migrations, they evolved in parallel in these different geographic regions, it being thought that “gene flow between the groups through interbreeding spread important changes throughout and was sufficient to maintain humans as a single species”.55 The Multiregional Evolution Model is not short of critics either, with Richard Klein stating:
“An obvious objection to multiregionalism is that it postulates substantial gene flow among small populations that were thinly scattered across three continents. In this light the multiregional model is not so much a theory as it is an after-the-fact explanation for proposed morphological resemblances between nonmodern and modern populations in Asia and Europe.”56
From an evolutionary point of view, it does not seem credible that for nearly two million years gene exchange between small Homo erectus populations, thinly scattered across three continents, helped keep them all united as a single species, while the different populations evolved separately into the anatomical modern human form, all getting there around the same time. With both the Out of Africa and Multiregional evolutionary models of human origins struggling to deal with the evidence, the biblical explanation of human origins appears much more plausible.
A biblical model
Regarding both the Out of Africa and Multiregional evolutionary models of human origins, as well as fossil humans like the Neanderthals and Homo erectus, creationist Carl Wieland has stated:
“I suggest that both contending evolutionary camps in this are each right about some things. There is a genetic continuity between us and these fossil humans, as both the fossil and nuclear DNA evidence shows. And we are all closely related, much closer in time, coming from one small population bottleneck (the Flood)—as the mtDNA evidence shows. Here, the biblical model would seem to provide the best of both worlds.”57
Creationists have different interpretations of Homo erectus fossils, although most models acknowledge them as being definitely human. One possible explanation, which I prefer, as to why ‘robust’ humans such as Homo erectus, Homo heidelbergensis and Neanderthals were different in morphology to anatomically modern humans, particularly in the skull, is that it could reflect changes in development of these early post-Flood individuals, compared to anatomically modern humans, possibly linked to longevity.58 It should be emphasized that I believe the key features of the robust morphology arose during ontogeny (i.e. development from fertilization of egg up to organism’s mature form). Whether this would have affected the time to reach maturation (i.e. attaining adult/mature form) is unclear, but this model does not require it to be different to extant humans. If the characteristic robust features were chiefly the result of developmental processes (genetically linked to greater longevity factors), then some of the robust fossils could be of individuals with the potential to live hundreds of years, and as such built robustly, but who died at a relatively young age (through illness, accident, conflict, etc.). If it was chiefly due to the aging process, every robust fossil discovered must, by default, have lived hundreds of years, which is unlikely; especially when some of the bony characteristics that distinguish these robust humans from moderns are already present in, for example, Neandertal specimens that are obviously still a long way from attaining adulthood, however long that may have taken.
Why would humans be designed more robustly when their lifespans were longer? Perhaps, having thickened cranial vault bones, a heavily built face, thick-boned jaws, and thick postcranial bones helped the body cope with long lifespans. If longevity was linked to development processes associated with robusticity, then robust features would be expected to disappear with shorter lifespans, although Wieland notes that “some of the genes coding for some of their distinctive bony anatomy were apparently passed on to some of today’s populations”.59 Hence, robust features do not correlate with the potential for longevity in recent times. Add environmental influences, as well as genetic effects like genetic drift, and you can possibly account for differences in skeletal features observed not only between early post-Flood humans, like Homo erectus, Homo heidelbergensis and the Neanderthals, but also between anatomically modern humans and these early robust post-Flood humans. As such, there would be regional variations in form in the past, as there is with people groups today.
Whether it is argued that skulls such as WLH 50 are ‘robust’ modern humans, Homo heidelbergensis, Homo erectus, or ‘archaic’ Homo sapiens, these are moot points, from the perspective of the position taken here, which is that Homo heidelbergensis and Homo erectus are not separate species either. Rather, the members allocated to these categories are considered to be fully human, descendants of Adam and Eve, and so should be included in Homo sapiens (though for the sake of identifying their categories their current names of classification were used).60
Conclusions
The Kow Swamp series crania are one group of ‘robust’ human fossils from Australia that were found to have Homo erectus-like features, but controversy later occurred after claims that some of the crania (e.g. Kow Swamp 5) had been artificially deformed. However, there are other crania that do not appear to be artificially deformed (e.g. Kow Swamp 1), and so this argument cannot be used against all the fossils. The Kow Swamp fossil remains were repatriated to the local (Echuca) Aboriginal community in 1990, and so are no longer available for study.
Concerning the robust WLH 50 skull, its large cranial capacity, estimated at about 1540 cc,41 would most likely have resulted in its arbitrary exclusion from Homo erectus, no matter where it had been found (an example of circular reasoning). However, the issue is not with attributing WLH 50 to a modern human, which is what it is, nor with seeing it as evidence of continuity with Ngandong Homo erectus populations. Rather, the problem is caused by not seeing Ngandong Homo erectus individuals as also modern humans (i.e. as Homo sapiens and descendants of Adam and Eve).
It clearly makes no sense to view the robust Willandra Lakes (e.g. WLH 50) and Ngandong individuals as separate species, but neither does it make sense to view some Homo erectus specimens (e.g. Ngandong) as modern humans, while viewing others (e.g. Sangiran or Zhoukoudian) as less evolved or subhuman.
If they are all considered as modern humans exhibiting a range of variation, i.e. as Homo sapiens and descendants of Adam and Eve, then there is no problem with the Willandra Lakes individuals being similar in morphology to the Ngandong individuals in Indonesia, as well as them being closely related. It is the belief in evolution, particularly the dominant ‘Out of Africa’ replacement model of human origins, that has caused the contradictions, as it cannot allow for robustly built humans, such as Homo erectus (or Homo heidelbergensis and Neanderthals for that matter), to be fully human.61
References and notes
- Campbell, B.G. and Loy, J.D., Humankind Emerging (8th edn), Allyn and Bacon, Needham Heights, MA, pp. 324–325, 2000. Return to text.
- Campbell and Loy, ref. 1, p. 325. Return to text.
- Campbell and Loy, ref. 1, p. 353. Return to text.
- Lieberman, D.E., The Story of the Human Body, Pantheon Books, New York, p. 72, 2013. Return to text.
- Antón, S.C., Homo erectus and related taxa, chap. 26; in: Begun, D.R. (Ed.), A Companion to Paleoanthropology, Wiley-Blackwell, West Sussex, UK, p. 497, 2013. Return to text.
- Stringer, C., Lone Survivors: how we came to be the only humans on earth, Time Books, New York, p. 15, 2012. Return to text.
- Bae, C.J., Douka, K., and Petraglia, M.D., On the origin of modern humans: Asian perspectives, Science 358( eaai9067), p. 3, 2017 | doi: 10.1126/science.aai9067. Return to text.
- Webb, S.G., The First Boat People, Cambridge University Press, Cambridge, UK, pp. 250–251, 274, 2006. Return to text.
- Thorne, A. and Wolpoff, M., Conflict over modern human origins, Search 22(5):177, July/August, 1991. Return to text.
- Thorne, A.G. and Macumber, P.G., Discoveries of Late Pleistocene Man at Know Swamp, Australia, Nature 238:316, 1972. Return to text.
- Durband, A.C. and Westaway, M.C., Perspectives on the Origins of Modern Australians (Chapter 4, pp. 123–150); in: F.H. Smith and J.C.M. Ahern (Eds.), The Origins of Modern Humans: Biology reconsidered, 2nd edn, John Wiley & Sons, Inc., Hoboken, NJ, p. 129, 2013. Return to text.
- Curnoe, D. and Thorne, A., The question of cranial robusticity, Before Farming, on-line version, 2006/2 Article 2, 2006. Return to text.
- Curnoe, D. and Thorne, A., Human origins in Australia: the skeletal evidence, Before Farming (on-line version) 2006/1 Article 5, 2006. Return to text.
- Durband, A.C., Artificial cranial deformation in Kow Swamp 1 and 5: A response to Curnoe (2007), HOMO—J. Comparative Human Biology 59:261–269, 2008. Return to text.
- Clark, J.L. et al., Identifying artificially deformed crania, International J. Osteoarchaeology 17:596, 605, 2007. Return to text.
- Clark, J.L. et al., Identifying Artificially Deformed Crania, International J. Osteoarchaeology 17:605, 2007. Return to text.
- Mulvaney, J. and Kamminga, J., Prehistory of Australia. Smithsonian Institution Press, Washington, p. 158, 1999. Return to text.
- Durband, A.C. and Westaway, M.C., Perspectives on the origins of modern Australians (Chapter 4, pp. 123–150); in: Smith, F.H. and Ahern, J.C.M. (Eds.), The Origins of Modern Humans: Biology Reconsidered, 2nd edn, John Wiley & Sons, Inc., Hoboken, NJ, p. 123, 2013. Return to text.
- Tenodi, V., Reconstructed face of a Stone Age woman unveiled in Greece would be forbidden in Australia, Pleistocene Coalition News 10(1):16, January–February, 2018. Return to text.
- Tenodi, V., Australian archaeological paradox: Did Homo erectus linger here? Pleistocene Coalition News 6(4):16–17, July–August, 2014. Return to text.
- Westaway, M., Human origins and the Mungo connection; in: Lawrence, H. (Ed.), Mungo Over Millennia: The Willandra landscape and its people, Maygog Publishing, Sandy Bay, Tasmania, pp. 24–30, 2006; p. 26. Return to text.
- Conroy, G.C. and Pontzer, H., Reconstructing Human Origins: A Modern Synthesis, 3rd edn, W.W. Norton & Company, New York, p. 592, 2012. Return to text.
- Durband and Westaway, ref. 18, pp. 128–130. Return to text.
- Westaway, ref. 21, p. 28. Return to text.
- Durband and Westaway, ref. 18, pp. 135–136, 138. Return to text.
- Hiscock, P., Archaeology of Ancient Australia, Routledge, London, p. 87, 2008. Return to text.
- Durband and Westaway, ref. 18, p. 138. Return to text.
- Conroy and Pontzer, ref. 22, pp. 592–594. Return to text.
- Conroy and Pontzer, ref. 22, p. 593. Return to text.
- Hiscock, P., ref. 26, pp. 93–94. Return to text.
- Hiscock, ref. 26, p. 94. Return to text.
- Flood, J., Archaeology of the Dreamtime: The story of prehistoric Australia and its people, rev. edn, Angus & Robertson, Sydney, pp. 40, 69, 1995. Return to text.
- Durband and Westaway, ref. 18, p. 133. Return to text.
- Grün, R., Spooner, N., Magee, J. et al, Stratigraphy and chronology of the WLH 50 human remains, Willandra Lakes World Heritage Area, Australia. J. Human Evolution 60:597, 604, 2011. Return to text.
- Hawks, J., Oh, S., Hunley, K. et al., An Australasian test of the recent African origin theory using the WLH-50 calvarium, J. Human Evolution 39:1–22, 2000. Return to text.
- Thorne, A.G. and Wolpoff, M.H., The multiregional evolution of humans, Scientific American 266:76–83, 1992. Return to text.
- Hawks, ref. 35, p. 21. Return to text.
- Webb, S.G., The First Boat People, Cambridge University Press, Cambridge, UK, p. 198, 2006. Return to text.
- Webb, ref. 38, pp. 194–208. Return to text.
- Webb, ref. 38, pp. 273–274. Return to text.
- Flood, J., ref. 32, p. 69. Return to text.
- Wolpoff, M.H., Hawks, J., Frayer, D.W., and Hunley, K., Modern human ancestry at the peripheries: a test of the Replacement Theory, Science 291:294, 2001. Return to text.
- Wolpoff et al., ref. 42, pp. 294–296. Return to text.
- Stringer, C. and Andrews, P., The Complete World of Human Evolution, 2nd edn, Thames & Hudson, London, pp. 142–143, 2011. Return to text.
- Barras, C., Ancient skull from China may rewrite our origins, New Scientist 236(3152):9, 18 November 2017. Return to text.
- Stringer and Andrews, ref. 44, p. 143. Return to text.
- Bae, C.J., Douka, K., and Petraglia, M.D., On the origin of modern humans: Asian perspectives, Science 358:eaai9067, pp. 3–4, 2017 | doi: 10.1126/science.aai9067. Return to text.
- Posth, C. et al., Deeply divergent archaic mitochondrial genome provides lower time boundary for African gene flow into Neanderthals, Nature Communications 8:16046, pp. 2, 6, 2017 | doi:10.1038/ncomms16046. Return to text.
- Gibbons, A., Neandertals mated early with modern humans, Science 56:14, 2017. Return to text.
- Posth et al., ref. 48, p. 5. Return to text.
- Hublin, J-J. et al., New fossils from Jebel Irhoud, Morocco and the pan-African origin of Homo sapiens, Nature 5 46:289–292, 2017. Richter, D. et al., The age of the hominin fossils from Jebel Irhoud, Morocco, and the origins of the Middle Stone Age, Nature 546:293–296, 2017. Return to text.
- Hershkovitz, I. et al., The earliest modern humans outside Africa, Science 359:456, 458, 2018. Return to text.
- Hershkovitz et al., ref. 52, p. 456. Return to text.
- Thorne, A.G. and Wolpoff, M.H., The multiregional evolution of humans, Scientific American (special edn: updated from the April 1992 edn.) 13(2):46–47, 2003. Return to text.
- Thorne and Wolpoff, ref. 54, pp. 46–47, 52. Return to text.
- Klein, R.G., The Human Career; Human biological and cultural, 3rd edn, The University of Chicago Press, Chicago, IL, p. 629, 2009. Return to text.
- Wieland, C., One Human Family: The Bible, science, race and culture, Creation Ministries International, GA, pp. 170–171, 2011. Return to text.
- Line, P., Explaining robust humans, J. Creation 27(3):64–71, 2013. Return to text.
- Wieland, C., Decreased lifespans: have we been looking in the right place? J. Creation 8:140, 1994. Return to text.
- Interestingly, some multiregionalists like Alan Thorne and Milford Wolpoff have, while they are evolutionists and believe that the robust features of e.g. Homo erectus were earlier than the gracile ones of Homo sapiens, also argued that specimens like Homo erectus should be renamed Homo sapiens. See: Wolpoff, M.H., Thorne, A.G., Jelinek, J., and Yinyun, Z., The Case for sinking Homo erectus, 100 years of Pithecanthropus is Enough!; in: Jens, L.F. (Ed.), 100 Years of Pithecanthropus: The Homo erectus Problem, Frankfurt am Main, Courier Forschungs-Institut Senckenberg 171:341–361, 1994. Return to text.
- We saw earlier that OoA proponents would also tend to oppose the idea that robust Australian skulls, like Kow Swamp 1 and WLH 50, are Homo erectus, because they ‘date’ as later on the evolutionary scale than some gracile Australian specimens, and also because they believe Homo erectus did not have the intelligence to make well-built watercraft and navigate the open seas, which would be required to reach Australia. Return to text.




Readers’ comments
Comments are automatically closed 14 days after publication.