

Journal of Creation 32(3):97–104, December 2018
Browse our latest digital issue Subscribe
Accounting for blighting plant and disfiguring animal diseases
One of the most significant groups of microbes causing plant disease are the fungi. Plant pathogenic fungi appear to have arisen primarily from those growing on decaying organic matter and from those capable of growing inside plants. Many of the species found inside plants either do not hinder growth or exert positive effects. In the natural environment, non-pathogenic fungi are subjected to changes in biological balance, stress, and alterations of the genetic fabric through mutations, faulty duplication events, limited gene transfer, and other phenomena. Such features appear to account generally for the emergence of pathogens from among previously non-pathogenic representatives. By contrast, few fungi are capable of causing mammalian disease. When they do, it is mainly as a result of gaining access through wounds, on account of environmental changes, or occurs when animals are weakened through stress or the immune system is down-regulated. The few fungi capable of causing disease in healthy individuals are infected mainly through inhalation of dust aerosols. It is difficult to imagine that dusty conditions existed in the Garden of Eden (Delight).
Many are aware of diseases of plants. For example, brown spots appear on the delicate rose blossoms as we are beginning to admire their beauty. Oranges begin to rot in our storage containers before we manage to eat them. Dirty black spots appear on the leaves of magnificent kangaroo paw plants. Fungal diseases of humans are relatively rare. However, those causing tinea or athlete’s foot and thrush (Candida) can be common.1 Some of the infections appearing on humans are disfiguring, such as basidiobolomycosis and paracoccidioidomycosis.
Many of the common diseases of plants are caused by fungi, the microscopic and classically thread-like organisms that are abundantly present in decaying organic debris in the soil. In a somewhat more complex form, we know them as mushrooms. A number of fungi form close relationships with plant roots and other living surfaces. Others actually penetrate plant cells or form networks of microscopic threads between the cells. Within this group of organisms, there is a developed ability to penetrate plants, which is significant to our discussion.
The useful and cooperative abilities displayed by fungi have been altered and fungi are now capable of causing many plant and some human diseases.
Fungal relationships with plants
The number of accepted fungal species is around 120,000,2 but only a small proportion of these are pathogenic. Those that are pathogenic are often closely related to non-pathogenic species. In fact, many of the genes identified in pathogenic fungi contribute to metabolic pathways found in all fungal species. Components of metabolic pathways regulating morphological and metabolic changes in response to stress and other external stimuli in non-pathogenic fungi are utilized in pathogenic fungi to regulate morphological and other changes associated with infection. In addition, those genes encoding enzymes capable of degrading plant cell walls or involved in forming infection structures are shared with saprophytes indicating a possible derivation from these fungi.3 However, there are several groups of proteins that are over-represented in plant pathogens that may have been the product of gene duplication and further changes.4 Certainly, variation in ploidy levels is known among selected plant and animal pathogens. Such changes may influence the expression of genes.5
The majority of fungi decompose organic debris (plant and otherwise) and live in close association with plant roots or on aerial plant surfaces. A surprising number of species have more intimate relationships with plants other than being pathogens. The mycorrhizal fungi (mutualists) invade plant roots to benefit their growth and confer resistance to pathogenic attack by select microbes (figure 1). Others grow mainly between cells without apparent effects on the operation of the host plant (endophytes). There is another group of fungi that grow on aerial surfaces (epiphytes) and live on the excretions and waste produced by sap-feeding insects.6 A small number of fungi among those found on aerial surfaces gain their nourishment from the plant through special intracellular feeding devices (haustoria) and are considered pathogens. While the compartmentalisation into categories, as suggested, is used for convenience of discussion, it should be understood that interactions between plants and microbes really operate on a continuum. Indeed, examples can be found where some mycorrhizal fungi can obtain some of their energy through the breakdown of organic matter and some wood-rotting fungi can form structures similar to those formed by genuine mycorrhizal fungi when they come in contact with roots.7
Many fungi infect plants, but do not produce symptoms. A considerable group found in this category are termed endophytes. Members typically live between the plant cells, but some may also penetrate cells. They may be highly localized in a plant or be systemic. It has been suggested that all plant species (vascular and non-vascular) possess such invaders and they may exceed a million species. When the plant is weakened by unfavourable climatic and other conditions, disease may develop. In favourable conditions, the endophytes have variable impacts on their hosts, but many appear to be beneficial. Those found in forage grasses may hasten seed germination, improve seed production and the defence and stress enduring capabilities of the plant, but the interactions are complex and depend very much on the strain of fungus and host involved. In some instances seed production is inhibited and infected plants resort to vegetative reproduction. In hosts other than forage grasses, additional benefits have been observed, such as improved nutrient uptake, expansion of the root system and subsequent increases in biomass and shoot growth, stimulation of the plant defences against pathogens, and increased stress tolerance.8
Whether an endophyte continues to grow without producing obvious symptoms in a plant depends on many factors including the developmental stage of both the host and fungus, the genetic capabilities of the fungus to cause disease and the plant to mount defences, and the environmental conditions prevailing. This means that plants placed under conditions of stress may be weakened, at which point the resident fungus may cause disease symptoms. For example, a strain of Leptosphaeria maculans isolated from the natural environment may subsequently cause disease in the same plant (Arabidopsis) under stressed laboratory conditions. There are a number of examples of changes in climatic conditions generating a switch to a pathogenic mode of operation where plant tissues are killed. Frequently endophytes become pathogenic following the senescence of tissues.9
The ability of fungi to live as an endophyte or mutualist involves eliciting a balanced response by the host plant to the presence of the organisms in its tissues. It is known that both endophytes and parasites use similar mechanisms of host recognition, but subsequent poorly understood biochemical features enable a friendly or unfriendly host response. It is known that a range of enzymes are released by endophytes that are capable of degrading organic components and aiding in infection. Why the enzymes and the toxins often produced do not cause disease is largely unknown, but the outcome seems to be related to the fine balance achieved between host defence reactions and fungus virulence factors.10 Potential pathogens among endophytes also may be held in check by special viruses that dampen their ability to be pathogens. Many of these viruses do not cause disease among the fungal isolates.11
In the classical evolutionary scheme of thinking, endophytes are commonly considered to have arisen from plant parasites.12 However, it is possible to conceive of an endophyte to parasite transition. I have adopted the latter scheme to account for some plant pathogens. This suggested pathway is based on the observed benefits often conferred on the host plant by endophytic microbial inhabitants. These benefits involve pathogen resistance, defence against herbivory, providing deterrents to seed predation, increasing plant competitive abilities and conferring stress resistance.13 Endophyte groups have family relatives that are well known degraders of organic debris, nematode-trapping fungi, and insect and plant pathogens (including rusts and smuts).14 Known pathogenic species have been identified as endophytes elsewhere, and as such do not cause any symptoms of infection.15 Indeed, one of the peculiarities of these organisms is that under certain circumstances endophytes may become pathogens or even vice versa, and a fungus endophytic with one plant may be pathogenic on another.10 Such changes commonly follow environmental stress.
The outcome of the interaction can be altered by endophytic and mycorrhizal fungi themselves being colonized by bacteria. These bacteria may have significant influences on the growth and fungal development before host infection takes place.16 One bacterium sometimes found in the hyphae of a mycorrhizal-like/endophytic fungus is Agrobacterium tumefaciens. The bacterium has growth-promoting influences and may function to enable the plant to be resistant to plant pathogens including epiphyte pathogens.17
Those fungi that operate regularly as plant pathogens release proteins into the environment, facilitating their ability to invade. The numbers of such proteins in the majority of pathogens is similar to that shown by fungi growing on organic debris. Those that are dedicated to keeping the host cells alive for the greatest period of time (e.g. obligate pathogens) generally have fewer secreted proteins to reflect their special relationship with plants, although there are exceptions.18
Obligate plant-inhabiting fungi
Some fungi are unable to live apart from plants and others have a limited existence outside plants. The former group includes organisms that damage (pathogens). Besides the pathogenic fungi of aerial plant parts such as rusts and mildews, there is a considerable group of beneficial mycorrhizal fungi that invade roots. Many are limited in their saprophytic ability and normally cannot complete their life cycle independent of plants. The mycorrhizal fungi usually confer benefits on the plants they colonise, but, under some conditions, they may become parasitic and reduce growth. Hence, again it is very much a case of maintaining a balanced relationship that determines whether an organism is considered a mutualist or a pathogen.7,19

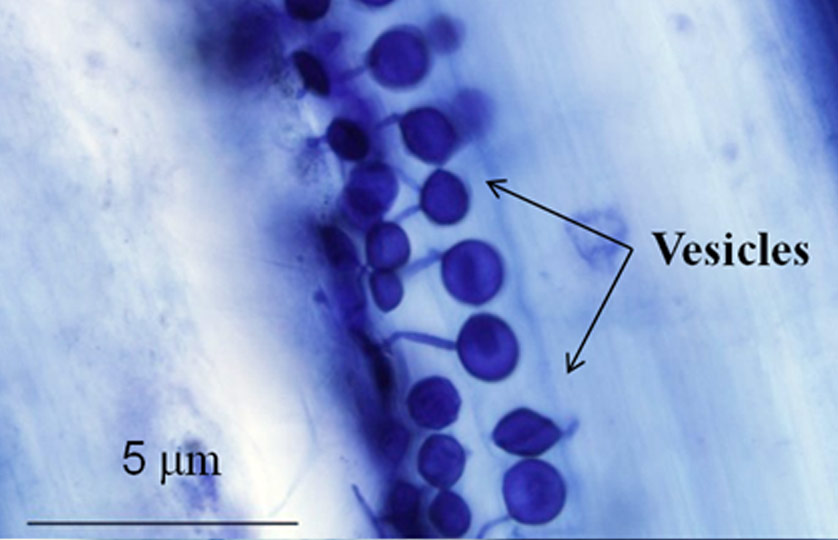
If we take very specialized groups of pathogens such as the powdery mildews and rusts, some interesting findings emerge. These organisms do not have an existence except as pathogens, thus they are termed obligate. In comparison to other fungi, they show a loss of certain biosynthetic pathways making it difficult, if not impossible, to culture them apart from a host plant. During the early stages of growth on a plant, some of the fungi (rusts) may be symptomless, essentially acting as endophytes. They gain their nourishment by growing intercellularly and producing special structures inside cells (haustoria).20 By contrast, the powdery mildews grow on aerial plant surfaces and obtain their nourishment intracellularly by means of haustoria (figure 2). It seems entirely possible for the organisms we now term obligate pathogens to have enjoyed a more balanced existence in the past where limited fungal growth in or on the plant led to sparse numbers of spores being formed. Conceivably, in the past, they might have possessed greater abilities for independent living, as genetic analysis can be taken to indicate. Their possession of intracellular feeding structures is shared by a small number of beneficial fungi (endophytes),9,18 hence strengthening the general argument in favour of a more balanced lifestyle in the past.
How this might have functioned is illustrated by two examples. The first is that systemic resistance to some powdery mildews is induced when a mycorrhizal-like fungus invades plant tissues. This only occurs when the invading fungus carries a specific bacterium (Agrobacterium) in its hyphae.17 The second example relates to a serious maize plant pathogen (Ustilago maydis), which does not fit into the obligate pathogen category, but nevertheless is not easy to cultivate. This organism can show symptoms in all the above-ground parts of the plant. If the major genes enabling colonization are silenced, then the fungus acts as an endophyte but is still able to complete its life cycle with the late production of spores.21
Plants possess complex defence mechanisms preventing invasion by unwanted microorganisms. Proteins are secreted by would-be pathogens. If these are recognized by resistant plants, then no invasion takes place, as the plant defences are activated. On the other hand, if the genes in the plant or microbe are altered, then the plant may be colonized by the microbe. Retrotransposons may give rise to changed proteins, which influence the microbe’s (including epiphytic species’) ability to invade.22
Plant and fungal genomes
Plants are designed to sense the presence of microbes that colonize them and this includes would-be pathogens. They have receptor molecules on cell surfaces that detect molecular patterns associated with microbes, which then stimulate the plant to mount a defence against the invader. Molecules released on account of microbial invasion also may stimulate defence mechanisms. The genetic systems of both plants and plant pathogens are finely balanced. The plant continually attempts to maintain its genetic integrity that enables it to be resistant, as through changing receptor characteristics. On the other hand, the microbe continually attempts to breach this barrier through such mechanisms as mutation, changes in the number and location of transposable elements, and loss of genes and other phenomena.18,23 The involvement of gene duplication and diversification being involved is strongly supported by the experimental data. Evidence for gene transfer is suggested mainly by comparative studies on gene sequences, but there are well-researched examples of bacterial DNA being integrated into host plant DNA (field) and bacterial DNA into yeasts and filamentous fungi (laboratory).24
Many of the plant pathogenic fungi sequenced have larger genomes than their closest non-pathogenic relatives. There are fungi with expanded genomes on account of possessing repetitive transposable elements, such as in Leptosphaeria maculans (which causes a stem disease in crucifers). Others appear to have lost genes and introns and have reduced transposon content. Some of the variations associated with pathogenic fitness may be generated through imprecision in the operation of repeat-induced point mutation machinery.3,25
The genome size of the pathogenic barley powdery mildew (Blumeria) again is larger than other related ascomycetes, but gene numbers are low in comparison to other fungal genomes. Genes commonly present in other filamentous fungi were absent in this and other powdery mildews. The number of transposable elements identified accounted for a sizeable 64% of the genome size. The increase in genome size and over-abundance of transposable elements (retrotransposons) appeared due to the absence of a repair pathway (repeat induced mutation), which also could be responsible for gene loss and reassortment of genetic information. It is postulated that the gene loss could have been responsible for these fungi becoming completely dependent on living cells for their existence. The mildew pathogens are among a group of fungi that avoid killing the plant cells. It is not surprising, then, that they produced a low number of secondary metabolites, which is a common feature in such pathogens. They also possessed a much reduced set of enzymes capable of degrading plant cell wall components.26
Single gene changes may enable a parasite to act as an endophyte and vice versa. This has been demonstrated experimentally with a pathogen of cucumbers (Colletotrichum magna) and an endophyte of ryegrass (Epichloë festucae). In the latter instance, mutation in a gene enabled the harmless endophyte colonist to cause severe plant stunting and death.10,27 It is also possible that gene changes in a pathogen may enable strong plant defences to be mounted, rendering the pathogen less effective in that role. In mutualistic fungi, underexpression of a particular gene may promote mycorrhizal infection while overexpression may impair it.18
The genetic information capable of enabling a fungus to cause disease may be carried by the organism, but may be silenced by other microbes associated with it. This has been rather convincingly shown with a soil-borne pathogen of lettuce. When closely associated bacteria were removed from the hyphae of a Fusarium fungus, it became pathogenic. This means that the associated bacteria were able to silence the expression of the fungal genes responsible for disease.28
Transfer of genetic information
Fungi are commonly filamentous organisms. The filaments are termed hyphae that originate often from vegetative cells, giving rise first to germ tubes and then hyphae. Those with the right genetic makeup may produce germ tubes that fuse (anastomose) to form a common structure that contains the genetic material possessed by both representatives in a common environment (hypha).
The significance of this phenomenon of anastomosis to form heterokaryons or dikaryons has been studied over a long period and confers abilities for adaptation on the new hypha produced as a result of both the fusion and subsequent events.29 The new hypha understandably contain nuclei from both contributing spores and may confer new abilities. One unusual consequence of such a fusion may be the appearance of enhanced disease expression, as illustrated in the following example. A disease of cereals (tan spot) that was first noticed around 80 years ago appears to be the result of transfer of a toxin gene (toxA) from one plant pathogen (Stagnospora nodorum on wheat) to a mild pathogen (Pyrenophora triticirepentis—yellow spot fungus on wheat) resulting in the emergence of a new and devastating disease. The original disease organisms can infect the same leaf, leaving the opportunity for hyphal fusion to take place. Although the relevant experiments have not yet been performed to show that fusion can occur between these two species, the DNA similarity between the species is strong and both are known to possess the capacity to anastomose.30 In related situations, it has been shown that whole chromosome transfer can occur between related fungi, under the right circumstances, even between those that are usually considered incompatible. Such transfer may enable non-pathogenic isolates of a particular fungus to become pathogenic. Hybrids may also form between different fungal species, when they are mixed in culture, to produce offspring that have an extended host range.31 Indeed, the transfer of material between seemingly incompatible fungi can be made possible by manipulating the chemical environment surrounding hyphae. Resulting anastomoses also allowed for the transmission of viruses between species of very different backgrounds.32
Genes and chromosome segments or even whole chromosomes may be transferred among fungal isolates and species via anastomoses involving hyphae or special protuberances arising from spores. Such fusions can occur on the host surface or in reproductive structures on the host.33
The transfer of plasmids may occur between parasitic fungi and their fungal host and between unrelated fungi that usually occupy the same ecological niche. Plasmid transfer between unrelated fungi is presumably through a temporary hyphal fusion.34 Of perhaps greater fascination is the transfer of DNA between bacteria and yeast (classical single-celled fungi) facilitated by conjugative plasmids. This appears to involve a conjugation process where a thin extension develops from the bacterium and fuses with the recipient yeast cell.35 It has been suggested, on the basis of gene homology, that toxin genes found in Photorhabdus (a bacterium symbiont in selected nematodes) have been involved in horizontal transfer to the endosymbiotic fungus Epichloë. In some bacteria, the toxin genes are plasmid-borne, making such a suggestion a real possibility.36 Mechanisms for such transfer have been identified in general terms and involve the formation of conjugation-type structures in interactions between bacteria and eukaryotes (plants, yeasts, filamentous fungi, and diatoms). Bacteria–plant transfers have been shown to occur in nature and bacterium-to-yeast transfer occurs in situations close to those encountered in the natural environment. In the plant-bacterium transfers, the donor DNA can be integrated into the recipient genome.24,37
Of interest also is the possible function of exosomes (cell-derived vesicles) in the transport of RNA. In powdery mildew infections (figure 2), exosomes may accumulate at contact sites with the plant. In other systems, micro-RNA components can be exchanged and induce gene silencing. With the mildew fungus Blumeria graminis, there is some evidence that trafficking of RNA occurs from the host to the fungus leading to silencing of genes in the fungus, which may translate into reduced haustorium formation. The phenomenon is being investigated as a means to control plant disease and has been shown to be successful with the rust fungus Puccinia tricticina.38 This mechanism potentially is highly significant in explaining the postulated altered interactions of powdery mildews and rusts since the beginning.
Intact yeast cells are also able to take up exogenous DNA (plasmid) via transformation. The plasmid DNA subsequently taken from the yeast was then capable of being reincorporated into bacteria. It is conceivable, but not proven, that conditions suitable for such transformation events could exist naturally in the environment.39 Transfer of plasmids between yeast species also has been shown experimentally, although the mechanism is not understood.40 This is an area of investigation that is worth watching closely.
The movement of genetic material from one organism to another can occur in the natural environment. This may be limited. Bacteria can be transformed in the natural environment by DNA from plants and other sources, if sufficient homology is present,41 and they in turn can donate bacterial DNA to fungi when the organisms are co-cultivated in the laboratory.42 The question might be asked whether there are bacteria present in soils that can act as a shuttle to pass genes between sources. The answer appears to be that such bacteria do exist, as illustrated below. A considerable group of fungi can form associations with and attachments to the soil bacterium Agrobacterium. In one well-executed study, attachment was confirmed by electron microscope. This enabled gene and plasmid transfer to take place and such transfer was confirmed by further analysis. Association between fungi and Agrobacterium in the soil environment and active gene transfer there is a possibility as this is the natural environment for both the bacterium and many fungi.43 Again, this is an area of investigation that might be followed closely, for it has significant implications. Readers might consider that bacteria other than Agrobacterium are capable of transferring DNA to plants, which can integrate into the genome and express its presence through bacterial enzyme activity.44
More distantly related to the transfer of genetic information is the phenomenon of one microbe invading another and enabling the consortium to become disease-producing. For example, a fungus (Rhizopus) pathogenic on rice seedlings has been shown to be invasive solely due to the presence of a bacterium (Burkholderia) in its hyphae. The bacterium produces a toxin which contributes significantly to the disease process.45
Fungal relationships with animals
The vast majority of fungi, and particularly those pathogenic to humans, can live a life independent of parasitism. They usually are harmless organisms found on or in the mammalian body or live on organic debris breaking it down into simpler forms. This means that changes in the animal host and environment are critical to their ability to infect.
A number of fungi are carried on or in the human body as harmless organisms, such as Candida albicans. If the biological balance existing between these organisms and the multitude of other microbes is upset, such as by antibiotic and chemotherapy treatments, illness, pregnancy, and other events, the organism can increase its population levels dramatically and cause disease. Among human beings, there are a number of developments that have occurred to account for increased host susceptibility and the upsurge in diseases caused by fungi. One significant factor is the large number of newly susceptible hosts produced by the burgeoning number of diabetic and AIDS patients.46
A second group of factors contributing to disease are adaptations in the microbe enabling it to accommodate to changing environments and become pathogenic. Virulence attributes (relative capacity to damage the host) can be thought of as factors that increase the ability of the organism to survive and colonize the mammalian host, but which are not essential for growth of the parasitic stage in vitro.47

The final group of factors that contributes to the emergence of disease is the existence of suitable environmental conditions. Virulent microbes and susceptible hosts may be in constant contact, but if the environmental conditions are unsuitable, disease will not develop. Good illustrations of this phenomenon are the common occurrence of Candida and Pneumocystis in the healthy human organism without causing disease. The environment may change when the immune system becomes less robust or the homeostatic status is altered through drug and other manipulations. It is then that disease develops.
A small number of fungi are able to cause disease in apparently healthy individuals (primary systemic pathogens—figure 3). These fungi enter the body through inhalation of aerosolized fragments of the organisms and their spores on account of disturbances in the environment. Other primary pathogens (not systemic but able to infect healthy people) may invade the subcutaneous tissues through a breach in the skin barrier.48 Species placed in the systemic group respond to the change of temperature in the mammalian body and switch from a filamentous to a yeast form of growth. This change enables the expression of genes that confer the ability to become pathogens in the animal body.49 In one pathogen (Histoplasma), the switch is regulated by a protein that is expressed at a higher level at animal body temperature.50 What role the genes responsible for such pathogenic ability play in the natural environment is not always understood. In the case of the pigment melanin or melanin-like pigments, which are produced by Histoplasma and other important human pathogens,51 their role is appreciated. In the natural environment melanin confers significant protection against damage to nucleic acid by ultraviolet light in a range of organisms.52 With Cryptococcus, capsule formation is a significant virulence determinant. In the natural environment, it appears the capsule protects the yeast from damaging bacteria.53
With the primary systemic pathogens, most depend for dispersal and intake into the mammalian body on the production of dust aerosols containing fungal elements, which are inhaled. In the pleasant environment described in the first two chapters of Genesis (a beautiful garden watered by a river and mist—2:6, 10), dust aerosols appear unlikely to have been produced. The human couple were to tend and keep God’s special creation (v. 15), not till and work the estate. However, once outside the Garden of Eden, and under the curse of sin, the soil was worked and thorns and thistles appeared (chapter 3:17–19), and the impact of climate change possibly was felt (Adam was destined to eat bread while sweating). Under these altered circumstances, organisms performing useful functions may have been given the opportunity to develop other abilities.
The appearance of pathogenic fungi on mammals is a result of changes in host, the microbe, and the environment. Stress and climatic changes, the occurrence of lifestyle diseases, accidents, complex surgical procedures, and imbalance observed in gene duplication and movement and other events all have played a part in the increasing significance of this group of microbes as pathogens.
Conclusion
When primary fungal pathogens, commensals, and opportunists are compared, there are few substantial differences in the features displayed. Each organism possesses a unique combination of factors (virulence composite) that enables it to be a successful colonist.54 The data available simply highlight the limitations in our knowledge and also illustrate that virulence genes vary in their significance depending on the strain and the host involved. A critical number of genes ultimately are necessary for the organism to cause disease. However, the gene-set necessary may change depending on the immune status of the host, on the site in the host encountered, and the form of the inoculum involved.55
The majority of fungi exist in the environment (terrestrial and aquatic) as decay agents of organic debris or form helpful associations with plants and animals. The change of activities from cooperative to destructive ones endangering life is ultimately a consequence of human disregard for God’s instructions and the search for an alternative philosophy of life. All nature has suffered the consequences. As a result of this deliberate choice by humanity, stress and dysfunction entered the world with widespread and unplanned destruction of life.
Several responses of fungi to these altered circumstances have been noted. Plants exposed to environmental and other stresses have been rendered variously susceptible to members of the fungal microbiota normally involved in the breakdown of organic debris. Those members devoted to living in harmony with plants or delivering benefits can also respond to the altered conditions by becoming pathogens. This transition can be easy, as the microbes already have well-versed systems that allow plant entry and a means of obtaining nutrients. In addition, plants and microbes are susceptible to changes in genome composition on account of mistakes in copying and movement of genes or gene segments within the genome.
Particularly with plant-invading fungi, changes to the genetic characteristics displayed can be accounted for by mutations, mismatch repair mistakes, parasexual phenomena, exchanges of genetic materials among fungi, or donations of DNA received from members of microbiota and others. The extent of acquisition of genetic information from alien sources is unknown, but there are well-established mechanisms for transfer including hyphal fusion, conjugation, and transformation.
The existence of obligate pathogens is the most difficult to explain. At least with one major group (powdery mildews) some useful information is available. They show a massive increase in the number of transposable elements, reassortment of genetic information with gene loss, and changes in transport of RNA from host to fungus. Such changes help to explain the current existence of a less-balanced relationship to the one that supposedly existed for this group of fungi in the distant past. The picture will undoubtedly clarify as more information is gathered.
Animals are susceptible to a small number of fungal pathogens. The majority enter through wounds and other portals made susceptible through environmental changes. The carriage of microbial spores and fragments in aerosols is an effective method of introducing unwelcome aliens into the body. As with plants, changes in both potential hosts and in the microbe may predispose to invasion and disease. The emergence of animal diseases can be explained, almost entirely, by invoking well-known nature- and human-induced phenomena.
References and notes
- Johnson, J., What we need to know about fungal infections, MedicalNewsToday, 17 June 2017, medicalnewstoday.com/articles/317970.php. Return to text.
- Hawksworth, D.L. and Lücking, R., Fungal diversity revisited: 2.2 to 3.8 million species, Microbiology Spectrum 5(4), 2017 | doi:10.1128/microbiolspec.FUNK-0052-2016. Return to text.
- Rouxel, T., Grandaubert, J., Hane, J.K. et al., Effector diversification within compartments of the Leptosphaeria maculans genome affected by repeat-induced point mutations, Nature Communications 2, article 202, 2011 | doi:10.1038/ncomms1189. Return to text.
- Soanes, D.N., Alam, I., Cornell, M. et al., Comparative genome analysis of filamentous fungi reveals gene family expansions associated with fungal pathogenesis, Plos One 3:1–15, 2008 | doi.org/10.1371/journal.pone.0002300 Return to text.
- Todd, R.T., Forche, A., and Selmecki, A., Ploidy variation in fungi—Polyploidy, aneuploidy, and genome evolution, Microbiology Spectrum 5(4), 2017 | doi:10.1128/microbiolspec.FUNK-0051-2016 Return to text.
- Chomnunti, P., Hongsanan, S., Aguirre-Hudson, B. et al., The sooty moulds, Fungal Diversity 66:1–36, 2014. Return to text.
- Koide, R.T., Sharda, J.N., Herr, J.R. et al., Ectomycorrhizal fungi and the biotrophy-saprotrophy continuum, New Phytologist 178(2):230–233, 2008. Return to text.
- Clay, K., Fungal endophytes of grasses, Annual Review of Ecology and Systematics 21:275–297, 1990; García-Guzmán, G. and Heil, M., Life histories of hosts and pathogens predict patterns in tropical fungal plant diseases, New Phytologist 201:1106–1120, 2014; Hamilton, C.E., Gundel, P.E., and Saikkonen, H.K., Endophytic mediation of reactive oxygen species and antioxidant activity in plants: a review, Fungal Diversity 54(1):1–10, 2012; Navarro-Melendez, A.L. and Heil, M., Symptomless endophytic fungi suppress endogenous levels of salicyclic acid and interact with the jasmonate-dependent indirect defense traits of their host, lima bean (Phaseolus lunatus), J. Chemical Ecology 40:816–825, 2014; Schulz, B. and Boyle, C., The endophytic continuum, Mycological Research 109(6):661–686, 2005. Return to text.
- DeLaye, L., García-Guzmán, G., and Heil, M., Endophytes versus biotrophic and necrotrophic pathogens—are fungal lifestyles evolutionarily stable traits? Fungal Diversity 60:125–135, 2013; Schulz and Boyle, ref. 8. Return to text.
- Kogel, K-H., Franken, P., and Hückelhoven, R., Endophyte or parasite—what decides? Current Opinion in Plant Biology 9(4):358–363, 2006; Schulz and Boyle, ref. 8. Return to text.
- Kobayashi, D.Y. and Hillman, B.I., Fungi, bacteria, and viruses as pathogens of the fungal community; in: Dighton, J., White, J.F., and Oudemans, P. (Eds.), The Fungal Community: Its organization and role in the ecosystem, 3rd edn, Taylor and Francis, Boca Raton, FL, pp. 399–422, 2005. Return to text.
- Clay, K. and Schardl, C., Evolutionary origins and ecological consequences of endophyte symbiosis with grasses, American Naturalist 160(suppl.):S99–S127, 2002. Return to text.
- Saikkonen, K., Faeth, S.H., and Helander, M. et al., Fungal endophytes: a continuum of interactions with host plants, Annual Review of Ecology and Systematics 29:319–343, 1998 | doi:10.1146/annurev.ecolsys.29.1.319. Return to text.
- Rodriguez, R.J., White Jr, J.F., Arnold, A.E. et al., Fungal endophytes: diversity and functional roles, New Phytologist 182(2):314–330, 2009. Return to text.
- Pan, J.J., Baumgarten, A.M., and May, G., Effects of host plant environment and Ustilago maydis infection on the fungal endophyte community of maize (Zea mays), New Phytologist 178(1):147–156, 2008. Return to text.
- Hoffman, M.T. and Arnold, A.E., Diverse bacteria inhabit living hyphae of phylogenetically diverse fungal endophytes, Applied and Environmental Microbiology 75(12):4063–4075, 2010. Return to text.
- Bonfante, P. and Anca, I-A., Plants, mycorrhizal fungi, and bacteria: a network of interactions, Annual Review of Microbiology 63:363–383, 2009. Return to text.
- Presti, L.L., Lanver, D., Schweizer, G. et al., Fungal effectors and plant susceptibility, Annual Review of Plant Biology 66:513–545, 2015. Return to text.
- Brundett, M., Diversity and classification of mycorrhizal associations, Biological Reviews 79(3):473–495, 2004. Return to text.
- Kemen, E. and Jones, J.D.G., Obligate biotroph parasitism: can we link genomes to lifestyle? Trends in Plant Science 17(8):448–457, 2012. Return to text.
- Brefort, T., Tanaka, S., Neidig, N. et al., Characterization of the largest effector gene cluster of Ustilago maydis, Plos Pathogens 10(7):e1003866, 2014 | doi:10.1371/journal.ppat.1003866. Return to text.
- Amselem, J., Vigouroux, M., Oberhaensli, S. et al., Evolution of the EKA family of powdery mildew avirulence-effector genes from the ORF 1 of a LINE retrotransposon, BMC Genomics 16:917, 2015 | doi: 10.1186/s12864-015-2185-x. Return to text.
- de Jonge, R., van Esse, H.P., Maruthachalam, K. et al., Tomato immune receptor Ve1 recognizes effector of multiple fungal pathogens uncovered by genome and RNA sequencing, PNAS 109(13):5110–5115, 2012; Dodds, P.N. and Rathjen, J.P., Plant immunity: towards an integrated view of plant-pathogen interactions, Nature Reviews Genetics 11(8):539–548, 2010; Stergiopoulos, I. and de Wit, P.J., Fungal effector proteins, Annual Review of Phytopathology 47:233–263, 2009. Return to text.
- Lacroix, B., Tzfira, T., Vainstein, A. et al., A case of promiscuity: Agrobacterium’s endless hunt for new partners, Trends in Genetics 22(1):29–37, 2006; Lacroix, B., Citovsky, V., Winans, S.C. et al., Transfer of DNA from bacteria to eukaryotes, mBio 7(4):e00863-16, 2016, mbio.asm.org/content/7/4/e00863-16.full. Return to text.
- Raffaele S. and Kamoun S., Genome evolution in filamentous plant pathogens: why bigger can be better, Nature Reviews Microbiology 10(6):417–430, 2012 | doi:10.1038/nrmicro2790. Return to text.
- Spanu, P.D., Abbott, J.C., Amselem, J. et al., Genome expansion and gene loss in powdery mildew fungi reveal tradeoffs in extreme parasitism, Science 330(6010):1543–1546, 2010. Return to text.
- Tanaka, A., Christensen, M.J., Takemoto, D. et al., Reactive oxygen species play a role in regulating a fungus–perennial ryegrass mutualistic interaction, Plant Cell 18(4):1052–1066, 2006. Return to text.
- Minerdi, D., Moretti, M., Gilardi, G. et al., Bacterial ectosymbionts and virulence silencing in a Fusarium oxysporum strain, Environmental Microbiology 10(7):1725–1741, 2008. Return to text.
- Shipton, W.A., The Biology of Fungi Impacting Human Health, Trafford, Singapore, pp. 94–96, 2012. Return to text.
- Friesen, T.L., Stukenbrock, E.H., Liu, Z. et al., Emergence of a new disease as a result of interspecific virulence gene transfer, Nature Genetics 38:953–956, 2006. Return to text.
- Ma, L.J., van der Does, H.C., Borkovich, K.A. et al., Comparative genomics reveals mobile pathogenicity chromosomes in Fusarium, Nature 464(7278):367–373, 2010; Mehrabi, R., Bahkali, A.H., Abd-Elsalam, K.A. et al., Horizontal gene and chromosome transfer in plant pathogenic fungi affecting host range, FEMS Microbiology Reviews 35(3):542–554, 2011. Return to text.
- Ikeda, K., Inoue, K., Kida, C. et al., Potentiation of mycovirus transmission by zinc compounds via attenuation of heterogenic incompatibility in Rosellinia necatrix, Applied and Environmental Microbiology 79(12):3684–3691, 2012. Return to text.
- Mehrabi et al., ref. 31. Return to text.
- Kempken, F., Horizontal transfer of a mitochondrial plasmid, Molecular and General Genetics 248(1):89–94, 1995. Return to text.
- Heinemann, J.A. and Sprague, G.F., Bacterial conjugative plasmids mobilize DNA transfer between bacteria and yeast, Nature 340(6230:205–209, 1989. Return to text.
- Han, J.E., Tang, K.F.J., Tran, L.H. et al., Photorhabdus insect-related (Pir) toxin-like genes in a plasmid of Vibrio parahaemolyticus, the causative agent of acute hepatopancreatic necrosis disease (AHPND) of shrimp, Diseases of Aquatic Organisms 113(1):33–40, 2015 | doi:10.3354/dao02830; Ambrose, K.V., Koppenhöfer A.M., and Belanger, F.C., Horizontal gene transfer of a bacterial insect toxin gene into the Epichloë fungal symbionts of grasses, Scientific Reports 4, article 5562, 2014 | doi:10.1038/srep05562. Return to text.
- Godio, R.P., Fouces, R., Gudina, E.J. et al., Agrobacterium tumefaciens-mediated transformation of the antitumor clavaric acid-producing basidiomycete Hypholoma sublateritium, Current Genetics 46(5): 287–294, 2004. Return to text.
- Nowara. D., Gay, A., Lacomme, C. et al., HIGS: host-induced gene silencing in obligate biotrophic fungal pathogen Blumeria graminis, Plant Cell 22(9):3130–3141, 2010; Valadi, H., Ekström, K., Bossios, A. et al., Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells, Nature Cell Biology 9(6):654–659, 2007; Panwar, V., McCallum, B., and Bakkeren, G., Endogenous silencing of Puccinia triticina pathogenicity genes through in planta-expressed sequences leads to the suppression of rust diseases on wheat, The Plant J. 73(3):521–532. 2013. Return to text.
- Nevoigt, E., Fassbender, A., and Stahl, U., Cells of the yeast Saccharomyces cerevisiae are transformable by DNA under non-artificial conditions, Yeast 16(12):1107–1110, 2000. Return to text.
- Mentel, M., Špírek, M., Jørck-Ramberg, D. et al., Transfer of genetic material between pathogenic and food-borne yeasts, Applied and Environmental Microbiology 72(7):5122–5125, 2006. Return to text.
- de Vries, J., Meier, P., and Wackernagel, W., The natural transformation of the soil bacteria Pseudomonas stutzeri and Acinetobacter sp. by transgenic plant DNA strictly depends on homologous sequences in the recipient cells, FEMS Microbiology Letters 195:211–215, 2001; Marcet-Houben, M. and Gabaldón, T., Acquisition of prokaryotic genes by fungal genomes, Trends in Genetics 26(1):5–8, 2009. Return to text.
- Sharma, K.S. and Kuhad, R.C., Genetic transformation of lignin degrading fungi facilitated by Agrobacterium tumefaciens, BMC Biotechnology 10:67, 2010 | doi: 10.1186/1472-6750-10-67 Return to text.
- Demanèche, S., Kay, E., Gourbière, F. et al., Natural transformation of Pseudomonas fluorescens and Agrobacterium tumefaciens in soil, Applied and Environmental Microbiology 67(6):2617–2621, 2001; Sharma and Kuhad, ref. 42. Return to text.
- Broothaerts, W., Mitchell, H.J., Weir, B. et al., Gene transfer to plants by diverse species of bacteria, Nature 433(7026):629–633, 2005. Return to text.
- Lackner, G., Möbius, N., Scherlach, K. et al., Global distribution and evolution of a toxinogenic Burkholderia-Rhizopus symbiosis, Applied and Environmental Microbiology 75(9):2982–2986, 2009; Partida-Martinez, L.P. and Hertweck, C., Pathogenic fungus harbours endosymbiotic bacteria for toxin production, Nature 437(7060):884–888, 2005. Return to text.
- Shipton, ref. 29, pp. 55–58. Return to text.
- Casadevall, A. and Pirofski, L., Host–pathogen interactions: the attributes of virulence. J. Infectious Diseases 184(3):337–344, 2001; Rappleye, C.A. and Goldman, W.E., Defining virulence genes in dimorphic fungi, Annual Review of Microbiology 60:281–303, 2006. Return to text.
- Reiss, E., Shadomy, H.J., and Marshall, G., Fundamental Medical Mycology, Wiley-Blackwell, Hoboken, NJ, pp. 14, 25–26, 2011. Return to text.
- Beyhan, S., Gutierrez, M., Voorhies, M. et al., A temperature-responsive network links cell shape and virulence traits in a primary fungal pathogen, PloS Biology 11(7):e1001614, 2013 | doi:10.1371/journal.pbio.1001614. Return to text.
- Nguyen, V.Q. and Sil, A., Temperature-induced switch to the pathogenic yeast form of Histoplasma capsulatum requires Ryp1, a conserved transcriptional regulator, PNAS 105(12):4880–4885, 2008. Return to text.
- Nosanchuk, J.D., Gómez, B.L., Youngchim, S. et al., Histoplasma capsulatum synthesizes melanin-like pigments in vitro and during mammalian infection, Infection and Immunity 70(9):5124–5131, 2002. Return to text.
- Gauslaa, Y. and Solhaug, K.A., Fungal melanins as a sun screen for symbiotic green algae in the lichen Lobaria pulmonaria, Oecologia 126:462–471, 2001; Ruan, L., Yu, Z., Fang, B. et al., Melanin pigment formation and increased UV resistance in Bacillus thuringiensis following high temperature induction, Systematic and Applied Microbiology 27:286–289, 2004; Shipton, ref. 29, pp. 129–131. Return to text.
- Abdulkareem, A.F., Lee, H.H., Ahmadi, M. et al., Fungal serotype-differences in bacterial-yeast interactions, Virulence 6(6):652–657, 2015 | doi:10.1080/21505594.2015.1066962. Return to text.
- Shipton, ref. 29, pp. 161–163. Return to text.
- Perfect, J.R., Genetic requirements for virulence in Cryptococcus neoformans; in; Domer J.E. and Kobayashi, G.S. (Eds.), The Mycota XII: Human fungal pathogens, Springer-Verlag, Heidelberg, pp. 89–112, 2004. Return to text.
- Zhang, T., Shi, N., Bai, D. et al., Arbuscular mycorrhizal fungi promote the growth of Ceratocarpus arenarius (Chenopodiaceae) with no enhancement of phosphorus nutrition, Plos One 7(9):e41151, 2012 | doi.org/10.1371/journal.pone.0041151. Return to text.




Readers’ comments
Comments are automatically closed 14 days after publication.