
The barrier has been breached!
Making a fool out of Professor Wise-man

A cardinal rule in biology is that the body cells and reproductive cells are separate in higher organisms.
This is called the Weismann barrier, after famous German evolutionist August Weismann (see Box) who discovered it. It was he who first realized the difference between the reproductive cells and the body cells in higher organisms.
This is not true in all species. The animals I studied for my PhD, corals, do not work like this. In corals, any cell can become a reproductive (i.e. ‘germ’) cell.1 For large coral colonies living in shallow water, this might include a cell from a lineage that has been blasted by high levels of mutation-inducing ultraviolet light for hundreds of years. How these animals have managed to persist over ‘evolutionary’ time is a bit of a mystery. Higher animals, however, carefully partition and protect their reproductive cells. They keep them in isolation and have various strategies for making sure they go through as few cellular divisions as possible.
In essence, the reproductive germ cells make the somatic (i.e. ‘body’) cells in order to make another generation of germ cells. This is a one-way system, with interesting parallels to the way workers in honeybee colonies only exist to ensure the queen bee can reproduce. The genetics of the worker bees is irrelevant because they do not pass on their DNA. Information cannot flow from the workers to the baby bees. In a similar way, information is not supposed to pass from the somatic cells to the germ cells. Only the information in germ cells should be passed on. This is the essential concept behind the Weismann barrier.
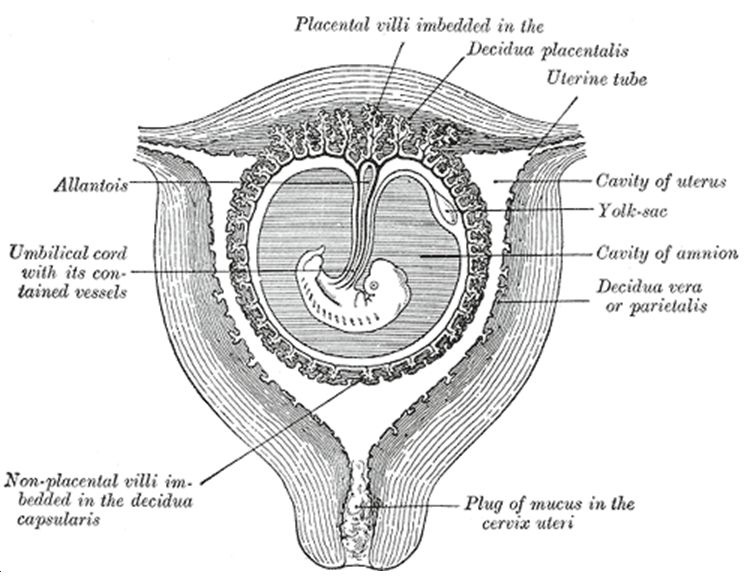
The reproductive cells are truly different. During the early stages of embryogenesis, they even spend time outside the embryo. The lineage of cells that are destined to become the germline will cluster on a structure called the allantois (figure 1).2 They hang out there for several days and reproduce several times. Later, they will form a cellular conga line, march toward the rapidly growing baby, get inside, and take up residence in the gonads.3 Prior to this, there is no morphological difference between male and female (there is always a genetic difference), but afterward the child will begin to develop secondary sex characteristics as the ovaries or testes start producing different levels of several hormones.4
There are also differences in the number of cell divisions between germline and somatic cells. This is important because polymerases (the cellular machines that copy DNA) are imperfect. They make about one mistake every billion letters or so, and the cell must copy six billion letters of DNA every time it divides.5 Thus, more cellular division equates to a higher mutation load. In the end, the eggs within a woman’s ovary are only separated from the original fertilized egg (at her conception) by about 24 cell divisions. The sperm-producing cells go through a longer maturation process. At puberty, they are about 30 cell divisions removed from the original zygote and will divide about 23 times per year after that. The final maturation steps require an additional five chromosomal replications.6 Yes, this means that fathers contribute more mutations than mothers.7 This is made even worse when you consider that eggs are held in protective custody for decades inside the ovary while spermatogonia constantly divide from puberty until the man’s death. This thought led to the development of an idea I call patriarchal drive, or the negative genetic effects of very old men having children in a small population. Being that he was the oldest father ever recorded, Noah was essentially ‘genetic poison’ to the post-Flood population. Likewise, the post-Flood patriarchs would have added significantly more mutations, per child, to the growing population than is typical today.8
One cannot underestimate how important Weismann’s concept was for the development of early evolutionary theory, namely that somatic and germline cells have different fates. In fact, Weismann was one of the first to strongly reject the Lamarckian view of inheritance (i.e. “the inheritance of acquired characteristics”). In doing so, he was going against Charles Darwin (1809–1882) himself. Darwin strongly held Lamarckian views. He believed the somatic cells budded off ‘corpuscles’ that migrated to the gonads. He called this pangenesis. Thus, Darwin believed information flowed from the body to the germ cells. In this way, traits that were acquired during life (stronger muscles, better eyesight, etc.) could be passed to the next generation. Weismann’s barrier completely contradicted this.

Together with the ‘central dogma of molecular biology’, the idea that information only flows in one direction (from DNA to RNA to protein), we have the two most important aspects of the gene-centric view of life. Richard Dawkins would never have written his famous book The Selfish Gene (discussed here) were it not for these ideas.
What if I told you that the Weismann barrier and the central dogma are both wrong? Yes, the ‘barrier’ has been breached. I hinted at this in an earlier article (see Genetic Diversity on Noah’s Ark), but new information has recently been discovered that makes it even more likely that Weismann was wrong.
Breaching the barrier
There are two interrelated areas where the Weismann barrier can be challenged. The first involves a temporary passing of epigenetic information from the adult to the baby. We have recently learned that sperm cells actively absorb RNA while they are being processed in the epididymis (a convoluted sperm-storage tube external to the testis).9 Vesicles are secreted from epididymal epithelial cells. These ‘epididymosomes’ contain various RNAs. Thus, there is a deliberately designed system to get somatic RNA into sperm cells. These will, in turn, be used by the early zygote as templates for protein translation, to control translation via RNA interference, and to store epigenetic tags that will turn different genes on or off. In other words, the environment of the father can influence which genes are active in the child. This invalidates the ‘gene centric’ view of life.
The second deals with a new discovery. It was already known that our cells actively turn RNA into DNA. The ‘central dogma’ claims the complete opposite. Our bodies produce an enzyme called reverse transcriptase that is designed to take RNA and turn it into DNA. This became well-known after the emergence of HIV. The RNA in this virus is regularly converted to DNA with our own reverse transcriptase and stored as DNA in the cell’s nucleus, making it very difficult to eradicate the virus because it can remain in latent form for decades. But we have just discovered an unexpected RNA to DNA pathway that involves an understudied DNA polymerase called Polθ (“pol theta”).
DNA polymerases do many things in our cells. They make copies of DNA and they turn DNA into many different forms of RNA. But we just learned that Polθ also works in reverse. In fact, Polθ is more effective at turning RNA into DNA than reverse transcriptase is. Polθ is involved in DNA repair. It uses an RNA template to determine what the DNA sequence was supposed to be before it got damaged.10 This might even be the explanation of a mystery from several years ago. Back in 2005, we learned that the plant Arabidopsis thaliana can fix its own mutations. This is even true when it is a homozygous recessive, in other words when the non-mutated gene is not even present in the plant. The authors titled their paper “Genome-wide non-mendelian inheritance of extra-genetic information … ”11 Considering everything said above, the phraseology they chose was shocking. This was, perhaps, just the tip of the iceberg. As more information like this comes in, especially now that we have a way to potentially explain such phenomena, neo-Darwinism will be weakened even further.
Yet, Polθ has no way of knowing how accurate the RNA template is. If the damaged gene is on the chromosome inherited from the father, it could be using an RNA coded by the gene inherited from the mother. It is theoretically possible that it might even be able to use an RNA template from a second but non-identical copy of the gene located somewhere else in the genome, or even an RNA template from an entirely different gene that happens to have a similar sequence in one region only. Even though this would violate the ‘central dogma’ that information only flows from DNA to RNA to protein, it is not necessarily a violation of the Weismann barrier. That is, unless the germ cells can use an RNA imported from the somatic cells, and we know germ cells (at least the sperm) actively absorb somatic RNA.
This new discovery opens up all sort of interesting new ideas. For example, what happens in the genome of a person with mixed-race ancestry? Is it possible that single letters from, for example, an “African” chromosome can be copied over onto a “European” chromosome? If a person has a maternal or paternal ancestor from a different racial group than all their other ancestors, what would be the fate of the mitochondrial genome or Y chromosome? Could Polθ take RNA that was absorbed from the body cells and use it to paper over mutations in those specific types of DNA? If so, the only source of RNA would be from a racially discordant set of chromosomes. If, for example, a person has an African paternal ancestor but for many generations everyone else in their family tree is European,12 the ‘mutations’ that accumulate could match those already found in Europe. The opposite would be true if, say, a person has European-type mitochondria but everyone else in the family tree is of African descent. This is not supposed to be true. Mutations are supposed to be random.
Also, what happens when environmental cues cause the production of specific RNAs? If the mother or father is starving, or sick, or happy and well fed, they would be expected to be producing different RNAs. These would serve as a source of both epigenetic regulation and as templates for Polθ DNA repair. Thus, the environment plays a possibly significant role in both the form of the adult organism and the rate and type of mutations that might accumulate in any lineage.
Most evolutionary speculations depend on oversimplified models of history. Historically, they have used the concept of a molecular clock (that mutations happen in a consistent and predictable rate over long periods of time), the ‘infinite alleles’ model (meaning all mutations are unique), and the idea that any mutation can be traced back to a specific ancestor (the activity of Polθ contradicts this). If the Weismann barrier has been breached, none of these things are necessarily true. Biology is suddenly a lot more complicated, and a lot more ‘messier’ as far as evolutionary modeling is concerned. They have been using overly simplified models to promote evolutionary theory. The true nature of biological complexity, however, means that the past century and more of evolutionary speculation can now be seriously questioned.
While the Weismann barrier still holds in general, we have no way of knowing how often it has been breached in history. This means we cannot easily put a timeframe on genetic events and we cannot necessarily pinpoint the geographic location of these events. For example, did humans evolve in Africa? There are multiple reasons for doubting this. Did Neanderthals split off from the early human lineage hundreds of thousands of years before Homo sapiens even appeared? This can no longer be supported from the models alone. The fall of the Weismann barrier is a seriously big deal.
Who was August Weismann?

August Friedrich Leopold Weismann (1834–1914) was an important early evolutionary theorist. Classically trained by his mother, an artist, and father, a university professor, he became a successful doctor and then professor of zoology at the University of Freiburg, where he founded the Zoological Institute in 1867. He predated the ‘rediscovery’ of Mendel’s work, so he did not have a complete theory of genetics, but he did reason that there was a complex, somewhat mysterious substance called ‘germ plasm’ that resided in the nuclei of germ cells. We now know this to be DNA. He also correctly reasoned that the germ plasm must be reduced by half before the egg and sperm unite. This happens during the process of meiosis, although he did not make the connection between germ plasm and what could be seen happening in chromosomes (meiosis was discovered by another German scientist, Oscar Hertwig, in 1876). It was Weismann who, in 1888, reported that he had cut the tails off over 900 mice for five generations, yet in the last generation the babies were born with normal-length tails. He, and others, reasoned that this disproved Lamarckian inheritance,13 although I think Darwin would not have appreciated the short timeframe of the experiment.

Wiseman may have been wrong, and he may have done damage to the Bible by promoting evolution, but he does not deserve as much disapprobation as some others. For example, his contemporary, also a German scientist, Ernst Haeckel (1834–1919), who was nicknamed “Darwin’s bulldog on the Continent”, deserves no mercy from historians. We called him an “apostle of deceit” for his fraudulent embryo diagrams, his fanciful descriptions of organisms that did not even exist, his promotion of mythical ape-like ancestors, and for his false claim (among many similar things) that there was a ‘fish-like’ stage of development in the human embryo.
Professor Weismann is quite possibly the inspiration of “Professor Wiseman”, a character in the book Curious George Gets a Medal (1957). This was written by the German-born Jewish couple Margaret (1906–1996) and H.A. (1898–1977) Reyersbach (later changed to “Rey”). In looks and intellect, the character mirrors Weismann in many ways. My mother read this book to me as a child and I, in turn, read it to my children. The subtle influence of evolutionary theory even percolates down into our children’s books. Yet, Professor Wiseman may not have been so wise after all.
References and notes
- This has recently been contradicted, but the results are far from conclusive. Either way, there is some point at which simpler organisms do not partition their germline and more complex organisms do. See Barfield, S., Aglyamova, G.V., and Matz, M.M., Evolutionary origins of germline segregation in Metazoa: evidence for a germ stem cell lineage in the coral Orbicella faveolata (Cnidaria, Anthozoa), Proc. Biol. Sci. 283(1822):20152128, 2016. Return to text.
- Figure 1 was taken from Gray, H., Anatomy of the Human Body, Lea and Febiger (Philadelphia and New York), 20th edition, 1918. This text is often just called Gray’s Anatomy and can be found in many modern versions. Dr Gray died of smallpox at the age of 34, after treating his 10-year-old nephew who had the disease and survived. Return to text.
- Wilhelm, D., Palmer, S., and Koopman, P., Sex determination and gonadal development in mammals, Physiol. Rev. 87(1):1–28, 2007. Return to text.
- Note: all babies do not “start out female”. This is an urban myth based on a gross misunderstanding of embryogenesis. I have also avoided a discussion of the complexities of sexual development. There is a lot more going on the embryo than just hormone production in the gonads. For example, the hypothalamus plays a significant role. See coggle.it/diagram/WvMd17PNQ5tdCKaV/t/sex-determination. Return to text.
- Yes, there are only 3 billion ‘letters’ in the human genome, but since humans are diploid, they carry two complete sets of chromosomes. Hence, our cells have to copy 6 billion letters every time they divide. Return to text.
- Crow, J.F., Age and sex differences on human mutation rates: an old problem with new complexities, J. Radiat. Res. 47(Suppl.):B75–B82, 2006. Return to text.
- Goriely, A. and Wilkie, O.M., Paternal age effect mutations and selfish spermatogonial selection: causes and consequences for human disease, Am. J. Hum Genet. 90:175–200, 2012. See also Jónsson, H. et al, Parental influence on human germline de novo mutations in 1,548 trios from Iceland, Nature 549:519–522, 2017. Return to text.
- Carter, R.W., Patriarchal drive in the early post-Flood population, J. Creation 33(1):110–118, 2019. Return to text.
- James, E.R. et al., The role of the epididymis and the contribution of epididymosomes to mammalian reproduction, Int. J. Mol. Sci. 21:5377, 2020. Return to text.
- Chandramouly, G. et al., Polθ reverse transcribes RNA and promotes RNA-templated DNA repair. Science Advances 7(24):eabf1771. See also the press release from Thomas Jefferson University “New discovery shows human cells can write RNA sequences into DNA”, phys.org/news/2021-06-discovery-human-cells-rna-sequences.html, 11 June 2021. Return to text.
- Lolle, S.J., Victor, J.L., Young, J.M., and Pruitt, R.E., Genome-wide non-mendelian inheritance of extra-genomic information in Arabidopsis, Nature 434:505–509, 2005. Return to text.
- E.g. King, T.E. et al., Africans in Yorkshire? The deepest-rooting clade of the Y phylogeny within an English genealogy, Eur J. Hum. Genet. 15:288–293, 2007. Return to text.
- Zou, Y., August Friedrich Leopold Weismann (1834-1914). Embryo Project Encyclopedia (2014-05-23). ISSN:1940-5030; embryo.asu.edu/handle/10776/7891. Return to text.




Readers’ comments
Comments are automatically closed 14 days after publication.