
An overview of the Denisovan puzzle

As ancient DNA analysis has become increasingly common, the identity of the mysterious Denisovans has gradually been revealed, through knowledge of their genome, as well as from more fossil bones being identified as belonging to this population. Although a complete picture is not yet available for the Denisovan puzzle, pieces are starting to fall into place. Here, an overview of this fascinating subject is given from a non-evolutionary viewpoint.
In 2008 a scrap of human finger bone was discovered in Denisova Cave (Figure 1), an isolated cave in the Altai mountains of southern Siberia.
A research team, led by Svante Pääbo, after extracting genetic material from the finger bone (Denisova 3; see Figure 2), sequenced its mitochondrial DNA (mtDNA) in 2010.1 The mtDNA analysis of the finger bone (a proximal fragment of a distal phalanx), said to be from a layer dated to between 48 and 30 ka (thousand years ago), found that Neanderthals and Denisovans differed from modern humans at an average of 202 and 385 nucleotide positions respectively.2 From this it was concluded that an “extinct species of human ancestor” had existed in Asia about 40 ka, and it was also “the first time a hominin has been described, not from the morphology of its fossilized bones, but from the sequence of its DNA.”3 Using the mtDNA from chimpanzees and bonobos as reference, the researchers estimated that at about 1 Ma (million years ago) “the new hominin’s mtDNA had shared a common ancestor with Neandertals and modern humans”.4

Long-age ‘dates’ herein, such as the above, are given to properly explain the research findings being reported on, but reflect the beliefs/assumptions within evolutionism, not those of the author (recent creation c. 6ka).
Later in 2010 the Pääbo research team published their analysis of the nuclear DNA of the Denisova 3 finger bone. They reported that the data suggested “it contributed 4–6% of its genetic material to the genomes of present-day Melanesians”, and designated the “hominin” population that it belonged to as Denisovans.5 They also stated that “the picture that emerges from analysis of the nuclear genome is one where the Denisova population is a sister group to Neanderthals.”6 On the analysis of the nuclear DNA Bustamante and Henn commented:
“… it seems that comparison of ancient and modern genomes processed at the same time provides a consistent picture of extra allele sharing between Denisovans and present-day Melanesians, as well as between Denisovans and Neanderthals.”7
The Pääbo team also reported on the morphology and mtDNA of a Denisovan upper molar tooth (Denisova 4; see Figure 3), its sequence said to differ at two positions from the mtDNA of the Denisova 3 finger bone, whereas it differed “at about 380 positions from both Neanderthal and present-day humans.”8 In 2015 the nuclear DNA sequence from Denisova 4 was analyzed, as well as the nuclear DNA and mtDNA sequence data from another Denisovan upper molar (Denisova 8) from Denisova Cave.9 According to the Pääbo team:

“The nuclear DNA sequences retrieved from Denisova 4 and Denisova 8 are more closely related to the Denisova 3 genome used to define the Denisovans as a hominin group than to present-day human or Neandertal genomes. Furthermore, the mtDNAs of the two molars form a clade with Denisova 3.”10
The Denisova 4 and Denisova 8 teeth were described as “very large compared with Neandertal and early modern human molars,” with the Denisova 8 molar said to be larger than the Denisova 4 molar.11 In 2017 the nuclear DNA and mtDNA sequence data of a deciduous lower molar from a fourth Denisovan individual (Denisova 2), also from Denisova Cave, was analyzed.12 From their analysis the Pääbo team concluded:
“On the basis of the number of “missing substitutions” in the mitochondrial DNA determined from the specimen, we find that Denisova 2 is substantially older than two of the other Denisovans, reinforcing the view that Denisovans were likely to have been present in the vicinity of Denisova Cave over an extended time period. We show that the level of nuclear DNA sequence diversity found among Denisovans is within the lower range of that of present-day human populations.”12
Up until the start of 2019 the above scrap of finger bone and three teeth represented the totality of fossil remains positively identified, via DNA analysis, as Denisovan. However, another finding of importance had also been revealed, after mtDNA and nuclear DNA analysis of a long bone fragment (Denisova 11) from Denisova Cave. Whilst mtDNA analysis in 2016 found the specimen to be carrying mtDNA of the Neanderthal type,13 even more interesting was the DNA analysis published in 2018, the latter finding that the individual was “a first-generation Neanderthal–Denisovan offspring”, i.e., the Denisova 11 bone fragment came “from an individual who had a Neanderthal mother and a Denisovan father.”14 For a catalogue of the supposed hominin remains (including non-Denisovans) from Denisova Cave up until the beginning of 2019 see Douka et al.15 In their paper these authors applied a Bayesian age modelling approach, combining “chronometric (radiocarbon, uranium series and optical ages), stratigraphic and genetic data to calculate probabilistically” the supposed age of the human fossils found in the Denisova Cave.16 As a result, Douka et al. concluded:
“Our modelled estimate for the age of the oldest Denisovan fossil suggests that this group was present at the site as early as 195,000 years ago (at 95.4% probability). All Neanderthal fossils—as well as Denisova 11, the daughter of a Neanderthal and a Denisovan—date to between 80,000 and 140,000 years ago. The youngest Denisovan dates to 52,000–76,000 years ago.”16
To add confusion to the identity of the Denisovans a recent genome sequencing study, analyzing “archaic introgression in a new dataset covering Island Southeast Asia (ISEA) and Papua”, suggested “considerable complexity in archaic contact, with modern humans interbreeding with multiple Denisovan groups that were geographically isolated from each other over deep evolutionary time.”17 As summed up by Dyani Lewis:
“In all, the team found DNA from three separate Denisovan populations: the Altai group, dubbed D0, and two others, D1 and D2. The latter two split off from the Altai Denisovans around 283,000 years ago, and from each other even further back, at 363,000 years ago.”18
Also, the “D1 population, whose sequences were especially enriched in people living on the island of New Guinea,” was said to be “as different from the other Denisovans as it is from Neanderthals,” and according to one of the researchers, probably warranted its own name.18
As if the supposed scenarios could not get more confusing, Ann Gibbons describes modeling indicating more potential mixing:
“And population geneticist Alan Rogers of the University of Utah in Salt Lake City said that his analysis suggests Denisovans and Neanderthals themselves were the product of interbreeding. He used computational modeling to test different scenarios for how Denisovans, Neanderthals, and modern humans acquired each other’s DNA. He concluded that the common ancestor of Denisovans and Neanderthals interbred with another extinct ‘superarchaic’ member of the human family, possibly H. erectus, about 700,000 years ago or so.”19
The above reference to a ‘superarchaic’ human appears to refer to a ‘ghost’ population, a population of which there is no “data from in unmixed form, but whose past existence can be detected from its genetic contributions to later people.”20 According to evolutionists there are at least three human ghost populations (Basal Eurasians, African Neanderthals and Ghost Denisovan), i.e., species or populations for which there is “no evidence except for their DNA”.21 However, one should remember that ancient DNA analysis, such as the above, is cluttered with unproven evolutionary assumptions and long ages, and so needs to be re-interpreted when integrated into a creation model.
Although confusing, one can conclude from the studies described above that the Denisovans were interbreeding with modern humans and Neanderthals. There is also strong evidence of Neanderthals interbreeding with modern humans.22,23 Between 2 and 4% of the genomes of non-Africans are estimated to be Neanderthal.24 Hence, by the biological species concept that makes them all part of the same species. Technically, they should all be classified as Homo sapiens. It also means that, from a biblical view, they were all members of the created human kind, and hence descendants of Adam and Eve.
Concerning the Neanderthals, a “species long maligned as knuckle-dragging brutes”, they are currently being “rehabilitated” by advocates such as the Portuguese archaeologist João Zilhão.25 Writing in the Smithsonian, Franz Lidz stated:
“Adaptation is key to Zilhão’s take on Neanderthals. He has long maintained that they were the mental equals of sapiens and sophisticated enough to imagine, innovate, absorb influences, reinvent them and incorporate that knowledge into their own culture. “Sure, there were physical differences between Neanderthals and modern humans,” he says. His tone is soft and measured, but there’s a flinty toughness to his words. “But Neanderthals were humans, and in terms of basic things that make us different, there was no difference.””26
But what about the Denisovans? What did they look like? A scrap of finger bone and three teeth tell next to nothing about what the Denisovans looked like. However, early in 2019 it was announced that a “palm-sized section of a braincase” was the first Denisovan skull fossil ever to be found.27 Discovered in Denisova Cave in 2016, the parietal bone fragment (Denisova 13) was attributed to being Denisovan based on its mtDNA, and consisted of two adjoining posterior half fragments of the left parietal.28 As described by Bruce Bower:
“Viola, of the University of Toronto, and his colleagues compared a digital reconstruction of the skull fragment with corresponding parts of 112 present-day human skulls and 30 Stone Age Homo skulls, including Homo sapiens and Neandertals. The Denisovan find didn’t fit neatly into any previously known Homo species. Some features linked the Denisovan fossil to Neandertals and to a 430,000-year-old Spanish Homo species that had Denisovan ancestry …. The Denisovan skull fragment is surprisingly thick, more like cranial bones of Stone Age Homo erectus, Viola said.”29
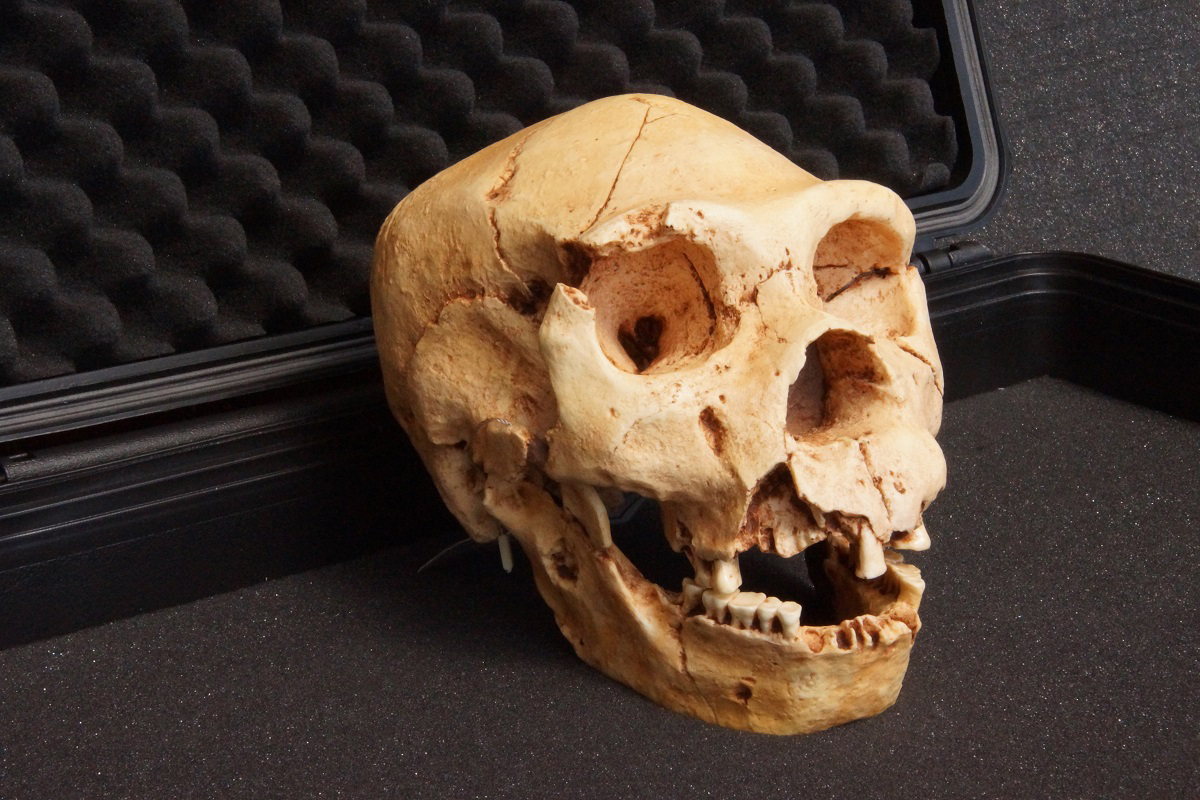
The Spanish Homo species mentioned above refers to the fossil remains from the Sima de los Huesos (SH) site in Sierra de Atapuerca, Burgos, Northern Spain. These remains (see Figure 4) are usually assigned to the species Homo heidelbergensis, as well as referred to as Middle Pleistocene hominins (or hominids). An almost complete mtDNA sequence of an SH specimen was reported in 2014 as “closely related to the lineage leading to mitochondrial genomes of Denisovans”.30 However, when the nuclear DNA was sequenced from two specimens in 2016, it was reported that the SH specimens “were related to Neanderthals rather than to Denisovans,” even though the mtDNA recovered from one of the specimens shared the earlier described relationship to Denisovan mtDNA.31
Evolutionists have presented various scenarios to explain the above finding,32 but they invariably lead to complex and unrealistic interbreeding storylines. However, if the evolutionary assumptions and long ages are dropped, then a simple explanation is that the Denisovan, Neanderthal and SH Homo heidelbergensis remains all belonged to populations that were fully human, and to various degrees interbred with each other.
That the Denisovan skull fragment was “surprisingly thick, more like cranial bones of Stone Age Homo erectus,” as described by Bower above, raises the question if the Denisovans were in fact Homo erectus.
In May 2019 Chen et al. reported on a Denisovan mandible (the right half), referred to as the Xiahe mandible (Figure 5), that had originally been found by a monk in 1980, at 3,280 m altitude, on the Tibetan Plateau in Baishiya Karst Cave, Xiahe county, China.33,34 The Xiahe mandible was identified as Denisovan by ancient protein analysis and said to be dated to at least 160 ka through U-series dating of carbonate matrix adhering to the mandible.35 The mandible, which has no chin, was described by paleoanthropologist Roberto Sáez:
“The mandibular morphology in general is primitive, short and very robust, and close to Homo erectus specimens. But the less elongated shape of the dental arcade is closer to that of the earliest Middle Pleistocene specimens of Neanderthals and Homo sapiens. The teeth are similar to the Denisovans’ from Altai mountains, and their morphology fits within Middle Pleistocene hominin variability.”36
Paleoanthropologist Jean-Jacques Hublin saw the jawbone, also the first known Denisovan fossil outside of Denisova Cave, as resembling the Penghu mandible found off the coast of Taiwan.37 The Penghu mandible is an “archaic Homo” specimen with many Homo erectus features, but it has not been assigned to a specific species within the genus Homo.38 Hublin is also quoted predicting “that most of the Chinese hominin fossil record younger than 350,000 years and older than 50,000 is made of Denisovans”.39

Whilst one cannot yet say with certainty that the Denisovans were Homo erectus, indications are that they were erectus-like, and whether you call them Homo erectus or Homo heidelbergensis (Middle Pleistocene hominins), is in many ways a moot point, as there appears to be little difference between them (see Figure 6). Paleoanthropologist Daniel Lieberman, an expert on the supposed evolution of the human head, commented that:
H. heidelbergensis and H. neanderthalensis crania are generally H. erectus-like but with a slight increase in face size and relative brain size. In contrast, H. sapiens represents a shift in craniofacial architecture, with a retracted, smaller face, and a more spherical vault.40
Here, fossils from human ‘species,’ such as Homo erectus, Homo heidelbergensis, and Homo neanderthalensis, are considered to have been early post-Flood humans, i.e., descendants of Adam and Eve, and so they should all be reclassified as Homo sapiens. Differences in morphological features between these human ‘species’ are considered as variations within the human kind, whether caused by genetic or environmental factors, and whether normal or pathological, that have nothing to do with ape-to-human evolution.

Some of the difference in the crania between Homo erectus, Homo heidelbergensis and Neanderthals may simply be due to differences in brain size, as brain growth largely determines the size and form of the neurocranium. Add environmental influences, along with genetic drift, and most of the differences between these early post-Flood humans can most likely be accounted for. Concerning why humans such as Homo erectus, Homo heidelbergensis and Neanderthals had differences in skull architecture compared to modern humans is open to debate, and not the focus herein, but one possibility is that it reflects differences in development of these early post-Flood individuals, possibly linked to longevity.41
A recent DNA study by Tucci et al., of a human pygmy population living in Rampasasa, Flores, near the Liang Bua cave where Homo floresiensis (aka the hobbit) was found, reported that the “genomes of Flores pygmies reveal a complex history of admixture with Denisovans and Neanderthals but no evidence for gene flow with other archaic hominins”, such as Homo floresiensis or Homo erectus.42 As stated by Gibbons, the “team found no trace of archaic DNA that could be from the hobbit.”43 However, the DNA of the hobbit is currently unknown, despite attempts to obtain it, so the study was making the unproven evolutionary assumption that Homo floresiensis was a different species to ‘modern’ humans and humans they categorize as Denisovans and Neanderthals. Perhaps there was no trace of the hobbit in the DNA of the Flores pygmies because Homo floresiensis never was a new or different species.
The tiny LB1 hobbit skull has similarities with humans categorized as Homo erectus, and a possible scenario is that it was a Homo erectus human with pathology.44 However, it is interesting that the Rampasasa pygmy population have traces of the mysterious Denisovans and Neanderthals in their genome. If the Denisovans were erectus-like people (the DNA of Homo erectus is also unknown), another possible scenario is that populations of ‘modern’ humans and Homo erectus (i.e., Denisovans) interbred on Flores, and individuals like the LB1 hobbit were the offspring, albeit with pathology.
The Denisovans are a fascinating subject. It is known from ancient DNA studies that the Denisovans interbred with ‘modern’ humans, and so by the biological species concept that makes them the same species, i.e., Homo sapiens. Hence, by logic the specimens in Homo erectus must also have been fully human, and part of the human kind, if they were Denisovan. The Denisovans appear to have been erectus-like, and it is hoped that further cranial bones of these humans will be identified, which should confirm whether they in fact were similar in morphology to humans classified as Homo erectus.
References and notes
- Dalton, R., Fossil finger points to new human species. Nature 464:472–473, 2010. Return to text.
- Krause, J. et al., The complete mitochondrial DNA genome of an unknown hominin from southern Siberia. Nature 464:894–895, 2010. Return to text.
- Brown, T.A., Stranger from Siberia, Nature 464:838, 2010. Return to text.
- Balter, M., Ancient DNA From Siberia Fingers a Possible New Human Lineage. Science 327:1566–1567, 2010. Return to text.
- Reich, D. et al., Genetic history of an archaic hominin group from Denisova Cave in Siberia. Nature 468:1053, 2010. Return to text.
- Reich, ref. 5, p. 1059. Return to text.
- Bustamante, C.D. and Henn, B.M., Shadows of early migrations. Nature 468:1045, 2010. Return to text.
- Reich, ref. 5, p. 1058. Return to text.
- Sawyer, S. et al. Nuclear and mitochondrial DNA sequences from two Denisovan individuals. Proceedings of the National Academy of Sciences of the USA, 112:15696–15700, 2015. Return to text.
- Sawyer, ref. 9, p. 15699. Return to text.
- Sawyer, ref. 9, p. 15697. Return to text.
- Slon, V. et al., A fourth Denisovan individual. Science Advances 3(7):e1700186, 2017. DOI: 10.1126/sciadv.1700186. Return to text.
- Brown, S. et al., Identification of a new hominin bone from Denisova Cave, Siberia using collagen fingerprinting and mitochondrial DNA analysis. Scientific Reports 6:23559, 2016. DOI: 10.1038/srep23559. Return to text.
- Slon, V. et al., The genome of the offspring of a Neanderthal mother and a Denisovan father. Nature 561:113, 2018. Return to text.
- Douka, K. et al., Age estimates for hominin fossils and the onset of the Upper Palaeolithic at Denisova Cave. Nature 565:640–644, 2019. For catalogue of supposed hominin specimens see Extended Data Fig. 1 and Supplementary Information pp. 27–34. Return to text.
- Douka, ref. 15, p. 640. Return to text.
- Jacobs, G.S. et al., Multiple Deeply Divergent Denisovan Ancestries in Papuans. Cell 177:1, 2019. https://doi.org/10.1016/j.cell.2019.02.035. Return to text.
- Lewis, D., DNA remnants of three separate Denisovan populations found in human genomes. 12 April 2019, https://cosmosmagazine.com/palaeontology/dna-remnants-of-three-separate-denisovan-populations-found-in-human-genomes. Return to text.
- Gibbons, A., Moderns said to mate with late-surviving Denisovans. Science 364:13, 2019. Return to text.
- Reich, D., Who We Are and How We Got Here, Pantheon Books, New York, p. 67, 2018. Return to text.
- Brahic, C., The ghosts within. New Scientist 240(3199):32, 13 October 2018. Return to text.
- Prüfer, K. et al., The complete genome sequence of a Neanderthal from the Altai Mountains. Nature 505:48, 2014. Return to text.
- Prüfer, K. et al., A high-coverage Neandertal genome from Vindija Cave in Croatia. Science 358:657, 2017. Return to text.
- Brahic, ref. 21, p. 30. Return to text.
- Lidz, F., The cave man cometh. Smithsonian 50(2):24–29, May 2019. Return to text.
- Lidz, ref. 25, p. 29. Return to text.
- Bower, B., The first known fossil of a Denisovan skull has been found in a Siberian cave. 29 March 2019, https://www.sciencenews.org/article/first-fossil-denisovan-skull-cave-siberia. Return to text.
- Viola, B.T. et al., A parietal fragment from Denisova cave. Abstracts of the 88th Annual Meeting of the American Association of Physical Anthropologists, March 27–30, 2019, p. 258. Return to text.
- Bower, B., The first known fossil of a Denisovan skull has been found in a Siberian cave. 29 March 2019, https://www.sciencenews.org/article/first-fossil-denisovan-skull-cave-siberia. Return to text.
- Meyer, M. et al., A mitochondrial genome sequence of a hominin from Sima de los Huesos. Nature 505:403, 2014. Return to text.
- Meyer, M. et al., Nuclear DNA sequences from the Middle Pleistocene Sima de los Huesos hominins. Nature 531:504, 2016. Return to text.
- Reich, D., Who We Are and How We Got Here, Pantheon Books, New York, pp. 71–74, 2018. Return to text.
- Chen, F. et al., A late Middle Pleistocene Denisovan mandible from the Tibetan Plateau. Nature 569:409–412, 2019. Return to text.
- Warren, M., Biggest Denisovan fossil yet spills ancient human’s secrets. Nature 569:16–17, 2019. Return to text.
- Chen, ref. 33, p. 409. Return to text.
- Sáez, R., Finally, the mandible of a Denisovan! 1 May 2019, https://nutcrackerman.com/2019/05/01/por-fin-mandibula-denisovano/ Return to text.
- Gibbons, A., Ancient jaw gives elusive Denisovans a face. Science 364:418–419, 2019. Return to text.
- Chang, C-H., et al., The first archaic Homo from Taiwan. Nature Communications 6:6037, pp. 1, 6, 2015. DOI: 10.1038/ncomms7037. Return to text.
- Wilson, C., Denisovans in Tibet. New Scientist 242(3228):8, 4 May 2019. Return to text.
- Lieberman, D.E. The Evolution of the Human Head, The Belknap Press of Harvard University Press, Cambridge, Massachusetts, p. 580, 2011. Return to text.
- Line, P., Explaining robust humans. Journal of Creation 27(3), 64–71, 2013. Return to text.
- Tucci, S. et al., Evolutionary history and adaptation of a human pygmy population of Flores Island, Indonesia. Science 361:511–512, 2018. Return to text.
- Gibbons, A., How islands shrink people. Science 361:439. 2018. Return to text.
- In an upcoming book I have a detailed chapter on Homo floresiensis. Return to text.




Readers’ comments
Comments are automatically closed 14 days after publication.