Ed. Note: this is the third instalment of a detailed critique of a major New Scientist anti-creationist diatribe (see introduction and index page). This one deals the widespread confusion between evolution and natural selection, actually a process discovered by creationists and an important part of the creation model.
Evolution: 24 myths and misconceptions
It will soon be 200 years since the birth of Charles Darwin and 150 years since the publication of On the Origin of Species, arguably the most important book ever written. In it, Darwin outlined an idea that many still find shocking – that all life on Earth, including human life, evolved through natural selection.
‘I have also seldom read a scientific book which makes such wide-ranging conclusions with so few facts supporting them. … Darwin wants to show that Arten [types, kinds, species] come from other Arten. I regard this as somewhat of a highhanded hypothesis, because he argues using unproven possibilities, without even naming a single example of the origin of a particular species.’ 1
And despite its hyped up ‘importance’, evolution provides no practical benefit to biology—see the detailed discussion in Does science need evolution? A modern evolutionist—and ardent misotheist—Jerry Coyne, argues that evolution is important as his (atheistic) theory of ‘How did we get here?’, but had to admit:
[I]f truth be told, evolution hasn’t yielded many practical or commercial benefits. Yes, bacteria evolve drug resistance, and yes, we must take countermeasures, but beyond that there is not much to say. Evolution cannot help us predict what new vaccines to manufacture because microbes evolve unpredictably. But hasn’t evolution helped guide animal and plant breeding? Not very much. Most improvement in crop plants and animals occurred long before we knew anything about evolution, and came about by people following the genetic principle of ‘like begets like’. Even now, as its practitioners admit, the field of quantitative genetics has been of little value in helping improve varieties. Future advances will almost certainly come from transgenics, which is not based on evolution at all.2
And even the claim about bacteria ‘evolving’ drug resistance is overstated, because this took evolutionists by surprise when it first occurred, and the changes involved are not those that would evolve bacteria into biologists. See the discussion in Anthrax and antibiotics: Is evolution relevant?
Darwin presented compelling evidence for evolution in On the Origin and, since his time, the case has become overwhelming.
Often those who declare the evidence to be ‘overwhelming’ or that ‘the debate is over’ say this to avoid debate. That’s why opposition is censored by ‘peer review‘ and opponents often demonized or discriminated against, as documented in the new film Expelled. As Thomas Sowell (1930– ) pointed out in another context (in his book Race and Culture about politically correct theories on race):
‘No belief can be refuted if it cannot be discussed.’
Countless fossil discoveries allow us to trace the evolution of today’s organisms from earlier forms.
Yet experts point out that it’s impossible to tell from fossils whether one creature was an ancestor of another, such as the late Colin Patterson. Furthermore, the fossil record should show gradual change from one kind of creature to another, millions of times over and it does not. Evolutionist Stephen Jay Gould called the scarcity of transitional fossils the ‘trade secret of paleontology’ (see this analysis as well as The Links Are Missing). For example, one evolutionist admitted:
‘The oldest bat fossils, belonging to an extinct lineage, were unearthed from rocks about 54 million years old, but the creatures that they represent aren’t dramatically different from living bats, says Mark S. Springer, an evolutionary biologist at the University of California, Riverside.
‘Hallmark features of these creatures include the elongated fingers that support the wing membranes and the extensive coiling of bony structures in the inner ears, a sign that they were capable of detecting the high-frequency chirps used in echolocation.’3
DNA sequencing has confirmed beyond any doubt that all living creatures share a common origin.
Ipse dixit (a dogmatic assertion without supporting evidence). All it can show are similarities; common origin as opposed to common design is an interpretation, and one fraught with problems. Rather, they support the biotic message theory, as proposed by Walter ReMine in The Biotic Message. That is, the evidence from nature points to a single designer, but with a pattern which thwarts evolutionary explanations because of the many similarities that cannot be explained by any theory of common ancestry—such as the incredible similarities between many marsupials and their placental counterparts (e.g. flying squirrels and flying phalangers—see Are look-alikes related?). Also, in most cultures that have ever existed, a consistent unifying pattern ‘brought honour to the Creator and would also indicate the Creator’s authority over and mastery of His creation.’4
Innumerable examples of evolution in action can be seen all around us, from the pollution-matching pepper moth to fast-changing viruses such as HIV and H5N1 bird flu.
Conflating natural selection and evolution is a staple of evolutionary propaganda. Recognizing this alone would almost be enough to see through the dogma. I’ll address these specific examples in later instalments when Le Page, the New Scientist author, cites them in more detail.
Evolution is as firmly established a scientific fact as the roundness of the Earth.
‘Evolution has been observed. It’s just that it hasn’t been observed while it’s happening.’5
And yet despite an ever-growing mountain of evidence, most people around the world are not taught the truth about evolution, if they are taught about it at all.
That’s true: the government schools and MMM (Mendacious Mainstream Media) teach evolution as fact, which is not the truth about it!
Even in the UK , the birthplace of Darwin with an educated and increasingly secular population, one recent poll suggests less than half the population accepts evolution.
So, despite the huge amount of evolutionary indoctrination, the indoctrinators are unhappy that it’s not working on everyone. And this includes even deliberately misleading students as long as it convinces them that evolution is true, since they believe, ‘Education is a subversive activity that is implicitly in place in order to counter the prevailing … deeply conservative religious culture.’
For those who have never had the opportunity to find out about biology or science, claims made by those who believe in supernatural alternatives to evolutionary theory can appear convincing. Meanwhile, even among those who accept evolution, misconceptions abound.
Yes, we have already encountered some propounded by Le Page, arguing that examples of an observable process (natural selection) is equivalent to proving the historical goo-to-you claim.
Most of us are happy to admit that we do not understand, say, string theory in physics,
True enough. Indeed, New Scientist itself has documented that even experts are confused by it,6,7 and reported the joke about why our universe is unique: it’s the only one string theory can’t explain. See also String theory unstrung.
yet we are all convinced we understand evolution. In fact, as biologists are discovering, its consequences can be stranger than we ever imagined. Evolution must be the best-known yet worst-understood of all scientific theories.
Then blame the propaganda pieces — like this one — that are more interested in point-scoring and word games than educating.
So here is New Scientist’s guide to some of the most common myths and misconceptions about evolution.
So here is New Scientist’s admission of ownership of this shoddy drivel, so they deserve all they get as a result. The gossip on the skeptics’ own websites suggested that Scientific American (SciAm) had suffered a financial downturn as a result of their ‘mistake’ in producing their anti-creationist article. A rebuttal on this site to a National Geographic anti-creationist tirade also resulted in people switching subscriptions to what is now our Journal of Creation.
There are already several good and comprehensive guides out there. But there can’t be too many.
Everything is an adaptation produced by natural selection
We tend to assume that all characteristics of plants and animals are adaptations that have arisen through natural selection. Many are neither adaptations nor the result of selection at all.
The 20-nanometer motor (height), ATP synthase (one nanometer is one thousand-millionth of a metre). These rotary motors in the membranes of mitochondria (the cell’s power houses) turn in response to proton flow (a positive electric current). Rotation of the motor converts ADP molecules plus phosphate into the cell’s fuel, ATP.
This is all true. But when it comes to the complex machinery of life, such as ATP synthase and the DNA winding motors, natural selection is the only game in town to try to avoid the overwhelming improbability of these machines arriving by chance (that is avoiding the abundantly clear implication that a super-intelligent Creator designed them).
This is a criticism of prominent evolutionary ‘adaptationism’. Gould and Lewontin invented the term ‘spandrel’ for features that supposedly arose not because of any direct adaptation, but as a by-product. This comes from cathedral architecture, a spandrel being the space between a rectangular corner and an inside curve such as an arch. It is also used of the space under a staircase. In cathedrals, this arch could be filled with a richly decorated panel or stained glass, and space under stairs is often used for a cabinet. But although the spaces could be put to use, the spaces were not intended per se, but were merely a consequence of something designed for another purpose (structural strength). Since artists use spandrels as a ‘canvas’ on which to paint their decorations, Gould argued that organisms could likewise use ‘functionless’ artefacts of anatomy for some new purpose.8
Take male nipples. Male mammals clearly don’t need them: they have them because females do and because it doesn’t cost much to grow a nipple. So there has been no pressure for the sexes to evolve separate developmental pathways and ‘switch off’ nipple growth in males.
We agree, except that it has nothing to do with ‘evolution’ switching or not switching as shown in Male nipples prove evolution?
…
Then there’s our sense of smell. Do you find the scent of roses overwhelming or do you struggle to detect it? Can you detect the distinctive odour that most people’s urine acquires after eating asparagus? People vary greatly when it comes to smell, largely due to chance mutations in the genes that code for the smell receptors rather than for adaptive reasons.
Certainly. So this has nothing to do with evolution, and it could be a built-in high-mutation system that can scan a wide range of chemicals. The elaborate design of the olfactory system, likely based on the principles of vibrational spectroscopy, would make this very easy, because the mutations could cause small changes in the quantum energy levels of the receptors. The elaborate olfactory system speaks of incredible design, not evolution.
Yet other features are the result of selection, but not for the trait in question. For instance, the short stature of pygmies could be a side effect of selection for early childbearing in populations where mortality is high, rather than an adaptation in itself.
That’s reasonable. But once again, nothing here is incompatible with the biblical Creation/Fall/Flood/Dispersion model, of which variation, natural selection and speciation are important parts.
Multiskilled genes
Another reason why apparent adaptations can be side effects of selection for other traits is that genes can have different roles at different times of development or in different parts of the body. So selection for one variant can have all sorts of seemingly unrelated effects.
A non-adaptive or detrimental gene variant can also spread rapidly through a population if it is on the same DNA strand as a highly beneficial variant. This is one reason why sex matters: when bits of DNA are swapped between chromosomes during sexual reproduction, good and bad variants can be split up.
Indeed, there is no dispute that sexual reproduction has its advantages. But explaining how this arose in the first place is a problem for evolutionists—see Evolution of sex?
Other features of plants and animals, such as the wings of ostriches, may once have been adaptations but are no longer needed for their original purpose.
There are at least three possibilities as to why ostriches, emus, etc have wings:
a) They derived from smaller birds that once could fly. This is possible in the creationist model. Loss of features is relatively easy by natural processes; acquisition of new characters, requiring new DNA information, is impossible.
b) The wings have a function. Some possible functions, depending on the species of flightless bird, are: balance while running, cooling in hot weather, warmth in cold weather, protection of the rib-cage in falls, mating rituals, scaring predators (I’ve seen emus run at perceived enemies of their chicks, mouth open and wings flapping), sheltering of chicks, etc. If the wings are useless, why are the muscles functional that allow these birds to move their wings?
c) It is a result of ‘design economy’ by the Creator. Humans use this with automobiles, for example. All models might have mounting points for air conditioning, power steering, etc. although not all have them. Likewise, all models tend to use the same wiring harness, although not all features are necessarily implemented in any one model. In using the same embryological blueprint for all birds, all birds will have wings.
Such ‘vestigial traits’ can persist because they are neutral, because they have taken on another function or because there hasn’t been enough evolution to eliminate them even though they have become disadvantageous.
Take the appendix. There are plenty of claims that it has this or that function but the evidence is clear: you are more likely to survive without an appendix than with one.
So why hasn’t it disappeared? Because evolution is a numbers game. The worldwide human population was tiny until a few thousand years ago, and people have few children with long periods between each generation. That means fewer chances for evolution to throw up mutations that would reduce the size of the appendix or eliminate it altogether — and fewer chances for those mutations to spread through populations by natural selection. Another possibility is that we are stuck in an evolutionary Catch-22 where, as the appendix shrinks, appendicitis becomes more likely, favouring its retention.
Yet in this supposed numbers game, there were enough mutations to cause bipedalism and development of a large brain enabling language development.
Wisdom teeth are another vestigial remnant. A smaller, weaker jaw allowed our ancestors to grow larger brains, but left less room for molars. Yet many of us still grow teeth for which there is no room, with potentially fatal consequences. One possible reason why wisdom teeth persist is that they usually appear after people reach reproductive age, meaning selection against them is weak.
This is also outdated. Wisdom teeth are rarely a problem for people, except those enjoying a modern western diet with soft, processed foods. This means less hard chewing during childhood jaw development, which causes a reduction in jaw size, and less stimulation of natural forward tooth movement in the jaw that would normally leave room for the third molar. Also, many dental surgeons caution against removal of these teeth unless they cause actual problems, not just as a preventive measure. See also Are wisdom teeth (third molars) vestiges of human evolution?
For all these reasons and more, we need to be sceptical of headline-grabbing claims about evolutionary explanations for different behaviours. Evolutionary psychology in particular is notorious for attempting to explain every aspect of behaviour, from gardening to rape, as an adaptation that arose when our ancestors lived on the African savannah.
Much change is due to random genetic drift rather than positive selection. It could be called the survival of the luckiest.
Take a look in the mirror. The face you see is rather different to that of a Neanderthal. Why? The unflattering answer could be for no other reason than random genetic drift. With features that can vary somewhat in form without greatly affecting function, such as the shape of the skull, chancemight play a bigger role in their evolution than natural selection.
Random genetic drift happens, but it has nothing to do with explaining how some reptiles changed into birds, for example. Such random changes do not explain the origin of the complex, integrated DNA coding necessary to specify how to make new features such as feathers. Neandertals were likely post-Babel humansadapted for the post-Flood Ice Age.
The DNA in all organisms is under constant attack from highly reactive chemicals and radiation, and errors are often made when it is copied. As a result, there are at least 100 new mutations in each human embryo, possibly far more. Some are harmful and are likely to be eliminated by natural selection — by death of the embryo, for instance. Most make no difference to our bodies, because most of our DNA is useless junk anyway.
More outdated nonsense, and largely derived from the evolutionary assumption that we have been around for millions of years. I.e. if much of our genome were functional, such a high rate of mutation would lead to error catastrophe unless most were non-functional. However, at least 97% of our DNA is now known to be transcribed, but much of it into regulatory RNA molecules rather than proteins. See Astonishing DNA complexity uncovered and update. This is further evidence that the evolutionary timescale is false because if we had been here for millions of years we would be extinct from the damage that mutations cause.
A few cause minor changes that are neither particularly harmful nor beneficial.
You might think that largely neutral mutations would remain restricted to a few individuals. In fact, while the vast majority of neutral mutations die out, a few spread throughout a population and thus become ‘fixed’. It is pure chance — some just happen to be passed on to more and more individuals in each generation.
Although the likelihood of any neutral mutation spreading by chance is tiny, the enormous number of mutations in each generation makes genetic drift a significant force. It’s a little like a lottery: the chance of winning is minuscule but because millions buy a ticket every week there is usually a winner.
And this drift has a good chance of eliminating even the rare beneficial mutations. This is a big problem for the gradualistic theories of evolution: the smaller the effect of a mutation, the more likely that drift will swamp its selective advantage. See this discussion in a review of Dawkins Climbing Mount Improbable.
As a result, most changes in the DNA of complex organisms over time are due to drift rather than selection, which is why biologists focus on sequences that are similar, or conserved, when they compare genomes. Natural selection will preserve sequences with vital functions, but the rest of the genome will change because of drift.
The actual evidence says the opposite. Most mutations have a small effect, so are immune from selection pressure. And genetic drift can often eliminate beneficial mutations.
See diagram (right) from Dr Sanford’s book (below)
Far more mutations are deleterious than advantageous. Individually, most have too small an effect to be acted upon by natural selection.
Drifting through bottlenecks
Genetic drift can even counteract natural selection. Many slightly beneficial mutations can be lost by chance, while mildly deleterious ones can spread and become fixed in a population. The smaller a population, the greater the role of genetic drift.
This is true. But then there is a lower supply of mutations, so it will take much longer for mutations to throw up anything useful.
Population bottlenecks can have the same effect. Imagine an island where most mice are plain but a few have stripes. If a volcanic eruption wipes out all of the plain mice, the island will be repopulated by striped mice. It’s a case of survival not of the fittest, but of the luckiest.
And nothing to do with evolution, because the disaster merely removed some information from the gene pool by chance.
Random genetic drift has certainly played a big role in human evolution. Human populations were tiny until around 10,000 years ago, and went through a major bottleneck around 2 million years ago. Other bottlenecks occurred when a few individuals migrated out of Africa around 60,000 years ago and colonised other regions.
There is no doubt that most of the genetic differences between humans and other apes — and between different human populations — are due to genetic drift. However, most of these mutations are in the nine-tenths of our genome that is junk, so they make no difference. The interesting question is which mutations affecting our bodies or behaviour have spread because of drift rather than selection, but this is far from clear.
Natural selection leads to ever-greater complexity
In fact, natural selection often leads to ever greater simplicity. And, in many cases, complexity may initially arise when selection is weak or absent.
If you don’t use it, you tend to lose it. Evolution often takes away rather than adding. For instance, cave fish lose their eyes, while parasites like tapeworms lose their guts.
Such simplification might be much more widespread than realised. Some apparently primitive creatures are turning out to be the descendants of more complex creatures rather than their ancestors. For instance, it appears the ancestor of brainless starfish and sea urchins had a brain.
Nevertheless, there is no doubt that evolution has produced more complex life-forms over the past four billion years. The tough question is: why? It is usually simply assumed to be the result of natural selection, but recently a few biologists studying our own bizarre and bloated genomes have challenged this idea.
No doubt? Well of course there is no doubt if you are a true believer and have decided that you don’t want to believe in a Creator; then evolution is the only game in town, by definition. See A tale of two fleas. And once again New Scientist promotes the outmoded idea that our genomes are full of junk (‘bloated’).
Rather than being driven by selection, they propose that complexity initially arises when selection is weak or absent. How could this be? Suppose an animal has a gene that carries out two different functions. If mutation results in some offspring getting two copies of this gene, these offspring won’t be any fitter as a result. In fact, they might be slightly less fit due to a double dose of the gene. In a large population where the selective pressure is strong, such mutations are likely to be eliminated. In smaller populations, where selective pressure is much weaker, these mutations could spread as a result of random genetic drift (see Natural selection is the only means of evolution) despite being slightly disadvantageous.
Gene or chromosome duplication is hardly the answer. In plants, but not in animals (possibly with rare exceptions), the doubling of all the chromosomes may result in an individual which can no longer interbreed with the parent type—this is called polyploidy. Although this may technically be called a new species, because of the reproductive isolation, no new information has been produced, just repetitious doubling of existing information. If a malfunction in a printing press caused a book to be printed with every page doubled, it would not be more informative than the proper book. (Brave students of evolutionary professors might like to ask whether they would get extra marks for handing in two copies of the same assignment.)
Duplication of a single chromosome (which contains many genes) is normally harmful, as in Down’s syndrome. Insertions are a very efficient way of completely destroying the functionality of existing genes, so if a duplicated gene is inserted randomly, it would likely cause damage to other functioning genes.
The evolutionist’s ‘gene duplication idea’ is that an existing gene may be doubled, and one copy does its normal work while the other copy is non-expressed. Therefore, it is free to mutate free of selection pressure (to get rid of it). However, such ‘neutral’ mutations are powerless to produce new genuine information. Dawkins and others point out that natural selection is the only possible naturalistic explanation for the immense design in nature (not a good one, as Spetner and others have shown). Dawkins and others propose that random changes produce a new function, then this redundant gene becomes expressed somehow and is fine-tuned under the natural selective process.
This ‘idea’ is just a lot of hand-waving. It relies on a chance copying event, genes somehow being switched off, randomly mutating to something approximating a new function, then being switched on again (how?) so natural selection can tune it.
Furthermore, mutations do not occur in just the duplicated gene; they occur throughout the genome. Consequently, all the deleterious mutations in the rest of the genome have to be eliminated by the death of the unfit. Selective mutations in the target duplicate gene are extremely rare—it might represent only 1 part in 30,000 of the genome of an animal. The larger the genome, the bigger the problem, because the larger the genome, the lower the mutation rate that the creature can sustain without error catastrophe; as a result, it takes even longer for any mutation to occur, let alone a desirable one, in the duplicated gene. There just has not been enough time, even with mythical evolutionary time, for such a naturalistic process to account for the amount of genetic information that we see in living things.
gene duplications are aberrations of cell division processes and are more likely to cause malformation or diseases rather than selective advantage
duplicated genes are usually silenced (no longer produce proteins) and subjected to degenerative mutations
regulation of supposedly duplicated gene clusters and gene families is irreducibly complex, and demands simultaneous development of fully functional multiple genes and switching networks, contrary to Darwinian gradualism.11
The more widely the duplicated genes spread in a population, the faster they will acquire mutations. A mutation in one copy might destroy its ability to carry out the first of the original gene’s two functions. Then the other copy might lose the ability to perform the second of the two functions. As before, these mutations won’t make the animals any fitter – such animals would still look and behave exactly the same – so they will not be selected for, but they could nevertheless spread by genetic drift.
This is another problem for the gene duplication idea.
Use your mutations
In this way, a species can go from having one gene with two functions to two genes that each carry out one function. This increase in complexity occurs not because of selection but despite it.
Once the genome is more complex, however, further mutations can make a creature’s body or behaviour more complex. For instance, having two separate genes means each can be switched on or off at different time or in different tissues. As soon as any beneficial mutations arise, natural selection will favour its spread.
If this picture is correct, it means that there are opposing forces at the heart of evolution. Complex structures and behaviour such as eyes and language are undoubtedly the product of natural selection.
Undoubtedly? Once again the philosophy of materialism reigns over rational thought and proper scepticism. Also, evolution does not explain the origin of eyes or language.
But when selection is strong—as in large populations—it blocks the random genomic changes that throw up this greater complexity in the first place.
Yet natural selection is the only real materialistic solution to the origin of complexity. That’s why atheists such as Dawkins defend(ed) it so strongly (see A Who’s Who of evolutionists).
This idea might even explain why evolution appears to speed up after environmental catastrophes such as asteroid impacts. Such events would slash the population size of species that survive, weakening selection and increasing the chances of greater genomic complexity arising through non-adaptive processes, paving the way for greater physical or behavioural complexity to arise through adaptive processes.
Sure, so lets improve the human race by exposing it to nuclear radiation, or chemical carcinogens that will speed up the mutation rate. This is the sort of fact-free story telling that the author has supposedly eschewed.
Evolution produces creatures perfectly adapted to their environment
You don’t have to be perfectly adapted to survive, you just have to be as well adapted as your competitors. The apparent perfection of plants and animals may be more a reflection of our poor imaginations than of reality.
It’s a theme repeated endlessly in wildlife documentaries. Again and again we are told how perfectly animals are adapted to their environment. It is, however, seldom true.
Take the UK ‘s red squirrel. It appeared perfectly well adapted to its environment. Until the grey squirrel arrived, that is, and proved itself rather better adapted to broadleaf forests thanks, in part, to its ability to digest acorns.
Certainly there is a gradation in complexity and a variety of features that enable plants and animals to adapt to different environments. But one would not claim that the Wright brother’s first man made (but not first overall) heavier-than-air flying machine was not designed, simply because there are far more complex planes now. A future instalment will discuss alleged design flaws further. But remember that the Bible tells us that we live in a fallen creation where things are no longer perfect.
There are many reasons why evolution does not produce ‘designs’ that are as good as they could be. Natural selection’s only criterion is that something works, not that it works as well as it might. Botched jobs are common, in fact. The classic example is the panda’s thumb, which it uses to grasp bamboo. ‘The panda’s true thumb is committed to another role. So the panda must… settle for an enlarged wrist bone and a somewhat clumsy, but quite workable, solution,’ wrote Stephen Jay Gould in 1978.
On closer inspection, however, there is nothing clumsy at all about the panda’s design.12 Instead, the ‘thumb’ is part of an elaborate and efficient grasping structure that enables the panda to quickly strip leaves from bamboo shoots.13
Claims that the panda’s thumb is some kind of non-designed ‘contraption’ is a smokescreen to distract from the real question—that evolution simply does not explain how life could start in a pond and finish with a panda.14
As this example shows, evolution is far more likely to reshape existing structures than to throw up novel ones. The lobed fins of early fish have turned into structures as diverse as wings, fins, hoofs and hands.
Yet the creatures they claim to be possible common tetrapod ancestors did not have five digits! Acanthostega had eight, while Ichthyostega had seven.
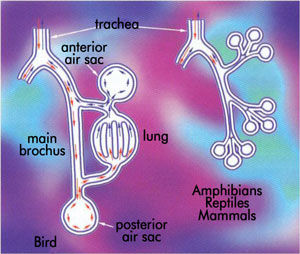
Credit Vij Sodera: One Small Step to Man (see below)Difference between reptile’s bellows lung and bird’s one-way lung. Click here to view larger image.
Many groups simply never evolve features that might have made them even more successful. Sharks lack the gas bladder that allows bony fish to control their buoyancy precisely, for example, and instead have to rely on swimming, buoyant fatty livers and, occasionally, a gulp of air. Similarly, mammals’ two-way lungs are far less efficient than birds’ one-way lungs.
But still good enough. On evolutionists’ dating, sharks have thrived for 300 million years without the gas bladder. One could ask why fish evolved such a thing that was clearly not necessary! And the transition from a bellows lung (as reptiles also possess) to an avian flow-through lung (see Blown away by design) has not been explained by evolutionists. One evolutionary expert in lungs explains the problem:
‘The earliest stages in the derivation of the avian abdominal airsac system from a diaphragmatic-ventilating ancestor would have necessitated selection for a diaphragmatic hernia [i.e. hole] in taxa transitional between theropods and birds.
‘Such a debilitating condition would have immediately compromised the entire pulmonary ventilatory apparatus and seems unlikely to have been of any selective advantage.’ 15
And sometimes creatures evolve features that actually reduce their overall fitness rather than increase it, such as the peacock’s tail.
Vij Sodera: One Small Step to Man (see below)Hypothetical stage of evolution of bird’s lung.
The complex design of the peacock tail is indeed a problem for evolutionists, especially as the sexual selection theory, which did not explain its intricate design anyway, has been disproven for this case—the very thing that Darwin invoked sexual selection to explain! See Peacock tail tale failure.
Use it or lose it
Continual mutation also means that if you don’t use it, you lose it. For instance, many primates cannot make vitamin C, because of a gene mutation. This mutation makes no difference to animals that get plenty of vitamin C in their diet. However, when the environment changes, such loss of function can make a big difference, as one primate discovered on long sea voyages.
Evolution’s lack of foresight can produce inherently flawed designs. The vertebrate eye – with its back-to-front wiring and blind spot where the wiring goes through the retina – is one example.
Humans are not running fast enough. Evolving through natural selection is about time and numbers. The number of new mutations that appear, and the number of chances that natural selection has to eliminate the harmful and favour the beneficial ones, depends on the size of a population, the number of offspring each individual has and on the number of generations, among other things.
We might like to think of ourselves as the most ‘highly evolved‘ species but, in terms of how many rounds of mutation and selection we’ve undergone, we are one of the least evolved species.
Around 10 billion new viral particles can be produced every day in the body of a person infected with HIV. By contrast, the total human population on Earth was no more than a few million until a few thousand years ago.
Indeed, Dr John Sanford (see above), shows that the known rate of harmful mutations accumulation really would have resulted in error catastrophe (i.e., extinction!) if we had really been around for millions of years16 (see his research papers published in secular journals17,18).
Furthermore, in a decade bacteria can produce 200,000 generations — about the number of generations of humans there have been since our lineage split from that of chimpanzees. So it’s hardly surprising that in less than a human lifespan we’ve seen the evolution of new diseases such as HIV and numerous antibiotic-resistant bacteria.
Behe’s second book, The Edge of Evolution,19 covers the issue of beneficial mutations and the limits of Darwinian processes. As his Ph.D. research involved malaria, he applies his expertise to the malarial parasite (Plasmodium falciparum) and the mutations humans have to deal with it, and the parasite’s counter-measures to human-made drugs.
One of the most effective anti-malarial drugs is chloroquine, because the parasite took longer to develop resistance to this. Behe shows that chloroquine resistance likely involves two specific mutations occurring together in the one gene. This explains why resistance to chloroquine took a long time to develop, whereas resistance to other anti-malarial drugs, which only needs one mutation, occurs within weeks. Behe works out the probability of this double mutation occurring in the same gene, using other scientists’ figures for the parasite’s population, etc.
If it took so much time for a double mutation to occur in an organism that has a huge population and short life cycle (and therefore huge opportunity for all manner of mutations to occur), then how long would it take for a double mutation to occur in an organism like a human, with a long generation time and small population? Behe showed that it would never occur even with evolutionary time assumed. And this is just one double mutation in a gene. So, any adaptation that requires two specific mutations in one gene to work, will never evolve in a human, and yet such must have happened numerous times if humans arose through evolutionary processes.
Behe also points out that the chloroquine-resistant parasites do worse than the non-resistant ones where there is no chloroquine. This suggests that the double mutation is informationally downhill, as usual. It seems that the reason that the parasite is resistant to chloroquine is that concentration in the parasite’s vacuole is reduced, and one mechanism is impaired uptake. According to one paper:
‘Chloroquine-resistant parasite isolates consistently have an import mechanism with a lower transport activity and a reduced affinity for chloroquine.’
This is the same principle that explains some antibiotic-resistant bacteria, where a mutation confers resistance by impairing a cell pump so the germ pumps in less of its would-be executioner.20
This leads to another of Behe’s major points: there is not so much an arms race as trench warfare or a scorched earth policy. Many of the changes are destroying machinery that the enemy could otherwise use. E.g. defenders will destroy their own bridges to prevent an enemy crossing, sabotage their own factories if the enemy is using them to churn out armaments, burn their own crops so the enemy will run out of food … This is why the world-class expert on sickle cell anemia, Dr Felix Konotey-Ahulu, rejects this icon of evolution .
Behe further reinforced the point by citing microbiologist Barry Hall on carbapenemeantibiotics:
‘Instead of assuming that [the chief kind of enzyme that might destroy these antibiotics] will evolve rapidly, it would be highly desirable to accurately predict their evolution in response to carbapeneme selection.’21
Hall showed that most antibiotics failed, but one (‘iminepen’) did not, simply because neither single nor double point mutations would suffice, but it would require more than two simultaneous mutations. Hall wrote that this was beyond the reach of mutation + selection:
‘The results predict, with >99.9% confidence, that even under intense selection the [enzyme] will not evolve to confer resistance to imipenem.’
Biophysicist Dr Lee Spetner in his book Not By Chance analyzes examples of mutational changes that evolutionists have claimed to have been increases in information, and shows that they are actually examples of loss of specificity, which means they involved loss of information (which is to be expected from information theory). See also this discussion, Is antibiotic resistance really due to increase in information?
Published: 5 December 2008
References
Interview/discussion with Professor Johann Blasius, 1859; reprinted in the German newspaper Braunschweiger Zeitung, on the 250th anniversary of the museum’s opening, 2004. Return to text.
Coyne, J., Selling Darwin (review of The Evolving World: Evolution in Everyday Life by David Minden, Nature 442:983–984, 31 August 2006 | doi:10.1038/442983a. Return to text.
Perkins, S., Learning to listen: How some vertebrates evolved biological sonar, Science News 167(20):314, 2005. Return to text.
Ideas needed—The hunt for a theory of everything is going nowhere fast, New Scientist 188(2529):5, 10 December 2005. Return to text.
Baffled in Brussels, New Scientist 188(2529):6, 10 December 2005. Return to text.
See Gould, S.J. and Lewontin, R.C., ‘The spandrels of San Marco and the Panglossian Paradigm: a critique of the adaptationist programme’, Proceedings of the Royal Society of London, Series B, 205(1161):581–598, 1979.Return to text.
Bollinger, R.R., Barbas, A.S., Bush, E.L., Lin, S.S. and Parker, W., Biofilms in the large bowel suggest an apparent function of the human vermiform appendix, J. Theor. Biol. 249(4):826–831, 2007. Return to text.
Scott, G.B., The primate caecum and appendix vermiformis: a comparative study, J. Anat. 131(3):549–563, 1980. Return to text.
Ruben, J.A., et al., Lung structure and ventilation in theropod dinosaurs and early birds, Science 278(5341):1267–1270, 14 November 1997. Return to text.
The new forward time genetics model Mendel’s Accountant demonstrates this, mendelsaccountaint.info; Baumgarder, J. et al., Mendel’s Accountant: A new population genetics simulation tool for studying mutation and natural selection; Sanford, J. et al., Using numerical simulation to test the validity of neo-Darwinian theory; in: Snelling, A., (ed.) Proc. 6th Int. Conf. Creationism, pp. 87–98, 165–175, Pittsburgh, PA, Creation Science Fellowship / ICR. Return to text.
Behe, M., The Edge of Evolution: The search for the limits of Darwinism, Free Press, NY, 2007; see review by Batten, D., Clarity and confusion, J. Creation 22(1):28–33, 2008. Return to text.
See Sarfati, J., Anthrax and antibiotics: Is evolution relevant? 2001–2005, creation.com/anthrax. Return to text.
Hall, B.G., In vitro evolution predicts that the IMP-1 metallo-beta-lacamase does not have the potential to evolve increased activity against imipenem, Antimicrob. Agents Chemother. 48:1032–1033, 2004. Return to text.
Comments are automatically closed 14 days after publication.
Feedback Guidelines
Be constructive & courteous. Don't attack individuals, denominations, or other organizations.
Stay on-topic. We're not here to debate matters like eschatology, baptism, or Bible translation.
Links to external sites and articles will be removed from your submission.
Privacy & Content Ownership
Comments become the property of Creation Ministries International upon submission and may be edited for brevity and clarity.
CMI may choose not to publish your comment depending on how well it fits the guidelines outlined above.
By submitting your comment you are agreeing to receive email updates from Creation Ministries International. You may unsubscribe at any time.
CMI records your real name, email address, and country as a sign of good faith. Privacy Policy
If your comment is published, your name will be displayed as ""
Cancel
Accept & Continue
Close
You are leaving CREATION.com
We have supplied this link to an article on an external website in good faith. But we cannot assume responsibility for, nor be taken as endorsing in any way, any other content or links on any such site. Even the article we are directing you to could, in principle, change without notice on sites we do not control.









Readers’ comments
Comments are automatically closed 14 days after publication.