

Journal of Creation 26(2):82–87, August 2012
Browse our latest digital issue Subscribe
Evaluating potential post-Flood boundaries with biostratigraphy—the Pliocene/Pleistocene boundary
Here I report a biostratigraphic analysis of 303 genera from 28 North American terrestrial mammalian families, in which all families contain members that are either extant or last appear in Pliocene or Pleistocene deposits. The distribution of these taxa within the Cenozoic rock record is used to evaluate proposed demarcations for the Flood/post-Flood boundary. A pronounced biostratigraphic break is expected at the Flood/post-Flood boundary since the final devastation and burial of pre-Flood nephesh creatures should be stratigraphically overlain by the arrival of post-Flood migrants. It is found that when the Flood/post-Flood boundary is placed at or near the Pliocene/Pleistocene boundary, then a significant number of mammalian genera (23%), and nearly every family (>96%), crosses this boundary. Because numerous boundary-crossing taxa would have to migrate from their North American pre-Flood habitats to board the Ark and return to their same continent of origin in the post-Flood world, it is unlikely that the Pliocene/Pleistocene boundary reflects the Flood/post-Flood boundary. Rather, the Flood/post-Flood boundary should be located below the Pliocene/Pleistocene boundary, at a geological location with a more pronounced biostratigraphic break.
The present distribution of living organisms is the consequence of numerous Flood-and post-Flood related events. For those nephesh creatures taken aboard Noah’s Ark, the biogeography of the modern species is primarily reflective of the migration and/or transport of original Ark-borne progenitor animals from the mountains of Ararat combined with intrabaraminic diversification (e.g. speciation). Whether baramins are best reflected at lower taxonomic levels (e.g. genera) or are more inclusive (families or higher taxonomic categories), intrabaraminic speciation events may be recorded in post-Flood sediments, leaving a fossil record of diversification within baramins leading to the present.
Evaluation of this record is the domain of biostratigraphy, the branch of paleontology dedicated to discovering the patterns of fossil occurrences within vertical sections of sedimentary rock. This may be done in local geological sections, or local sections can be correlated and combined for broad-scale evaluations (regional, continental, and global). Biostratigraphy also falls within the broader discipline of geological correlation, the process of linking/matching rock units over distances in which they are not seen.
While a number of criteria have been offered for determining the location of the Flood/post-Flood boundary, there remains as yet no consensus. Opinion is primarily (though not entirely) split between a boundary at or near the Cretaceous/Paleogene (= Tertiary)1–3 or the Pliocene/Pleistocene4,5 divisions. I submit that a robust biostratigraphic analysis aids in determining the location of the Flood/post-Flood boundary, since a pronounced biostratigraphic break marking the termination of the Flood should be expected by all creationists. The reasons are as follows.
- Since the pre-Flood distribution of continents was markedly different than the modern distribution,1,6 pre-Flood ecosystems from any given continent would be significantly different from those found on that same continent after the Flood.
- Even if the continental configuration was identical to today (which is unlikely), the majority of young-earth creationists maintain that Earth’s climate was more equitable, particularly in (presently) temperate and polar regions. Hence pre-Flood organisms borne by the Ark would face differing climates upon their return to the same continent and latitude. Such climatic differences would only become more pronounced leading up to and throughout the post-Flood Ice Age7 (which, in accordance with ‘high’ boundary placement, occurs very shortly after the end of the Flood).
- As a result, it is unlikely that the post-Flood dissemination of animals would result in a return to their pre-Flood geographic locales. In other words, it is unlikely that species of baramins taken aboard the Ark would display a proclivity to migrate to the graveyards of their deceased, pre-Flood baraminic kin.
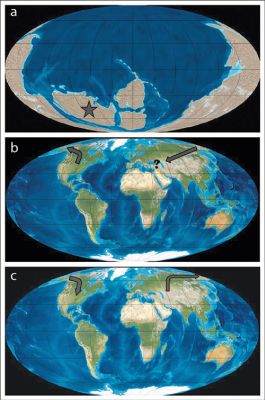
Figures 1a–c illustrate the biogeographic problem at the heart of the Flood/post-Flood boundary debate. Figure 1a represents the starting point of travel that a representative of a North American mammal baramin needs to make, given a Rodinia-type continental configuration prior to the Flood. This represents the current, conventionally inferred positions of the continents just prior to the Cambrian, and stands in accord with substantial tectonic reorganization during Noah’s Flood, such as those envisioned in Catastrophic Plate Tectonics. Figure 1b depicts the travel route from a modern plate configuration, which is nearly identical to the continental configuration at the Pliocene/Pleistocene boundary (the question mark reflects that the location of Noah’s Ark prior to the Flood is unknown; it may have been in Asia, or elsewhere). This view requires that significant lateral tectonics did not occur during Noah’s Flood. Again, I believe this second option is incorrect, but it is provided here for illustrative purposes. Once the Flood abates, the Ark-borne progenitors and/or their offspring must migrate to North America (figure 1c).
Regardless of the initial continental configuration, the following must be true if the Flood/post-Flood boundary is placed at or near the Pliocene/Pleistocene boundary: North American fossil taxa must have either inhabited the locations of their present fossil deposits (in North America) prior to the Flood, or they must have been transported exceptionally long distances en masse to North America during the Flood. From the dissemination of nephesh kinds released from the Ark, one would also expect that the post-Flood baramin representatives would follow a ‘sweepstakes’ pattern of opportunistic migration and inhabitation of the new post-Flood world and its varied environments. Given this and the above-mentioned differences in climate (especially pronounced given the location of North America/Laurentia in figure 1a), there would likely be no preference for any particular baramins to migrate back to the starting locations of their now-deceased, pre-Flood kin.
But what if the Flood/post-Flood boundary is not/ cannot be placed at a particular geologic location? That is, what if the Flood ended in one location that geologists call ‘Eocene’ and elsewhere in the ‘Pliocene’? While this may be possible for certain deposits (e.g. the marine sediments of the southeastern United States display a marine → terrestrial transition reflective of continual sea level drop through much of the Cenozoic; in this case, some areas may be post-Flood earlier than other, still-inundated areas), it is unlikely to apply here. In North America, the vast majority of the mammals evaluated here are found in sedimentary deposits from regionally restricted terrestrial basins, rather than trans-continental sedimentary deposits likely to be formed under Flood condition. This fact was part of the rationale for placing the end of the Flood near the Cretaceous-Paleogene (= Tertiary) boundary by Austin et al.1 in their description of Catastrophic Plate Tectonics.
Methods
Selection of North American mammals
Ideal groups to test the argument for a Pliocene/ Pleistocene location for the Flood/post-Flood boundary are North American mammals. These groups benefit from a long history of intensive, well-documented collection and study. There are dozens of notable localities with excellent stratigraphic sections throughout the Cenozoic from far-ranging locations across the continent (ranging from California and Oregon to Nebraska, South Dakota, and Florida). Furthermore, two recent publications8,9 provide a comprehensive treatment of the diversity, biogeography, and biostratigraphy of the entire North American Cenozoic mammal fossil record, and these data have been imported into searchable online databases.10
Creation of biostratigraphic range charts
North American mammalian families were tabulated and analyzed using the Paleobiology Database.10 The Paleobiology Database is a collaborative online repository of paleontological information. In particular, the Paleobiology Database includes taxonomic, biogeographic, and biostratigraphic data for fossils described in the professional paleontological literature and curated in museums and university repositories. This database is searchable using a variety of browser-based tools and methods, and permits downloading of data for additional methods by researchers.11
For this analysis, mammalian families were chosen from the following Orders: Artiodactyla, Carnivora, Edentata/Xenarthra, Insectivora, Lagomorpha, Marsupialia, Perissodactyla, and Proboscidia. Each of the families selected from these orders contain genera that are either extant or cross the Pliocene/Pleistocene boundary, and therefore exist until the Ice Age, which is recognized by creationists to be a post-Flood event.7

To conduct the analysis, I employed the following methods:
- From the main page at www.pbdb.org, the ‘Count taxa’ tool from the ‘Analyze’ tab was selected to create a list of the fossil genera within each queried family. I entered the family name (e.g. Canidae) into the ‘Taxonomic group’ box. I then clicked ‘Submit Query’, and the tool produced a list of genera and species.
- I copied and imported this list of genera into the ‘Analyze taxonomic ranges’ tool, which builds biostratigraphic range charts from known fossil occurrences, generated from the published literature and records from museum collections. After entering the genus list, I selected ‘as entered’ in the ‘Break taxa into’ box and submitted the query.
- The next page which loads is the ‘Confidence interval options’ page. Under ‘Time scale’, researchers can choose among a number of biostratigraphic methods for graphical output. I selected ‘North American Land Mammal Ages’ (see below for discussion).
- On this same page, I selected ‘no confidence intervals’ under the ‘Estimate’ box. This retrieves only the raw occurrence data, with no statistical estimates on biostratigraphic ranges above or below documented first and last appearances.
- For visual ease of evaluation, I selected ‘last appearance’ for the ‘Order taxa by’ box and submitted the query.
Standard geological chronology vs North American land mammal ages
I evaluated the differences in reporting which result from selecting two timescales applicable to the Cenozoic mammal record of North America: stages (e.g. Eocene, Oligocene, Miocene) and North American Land Mammal Ages (herein NALMAs). The NALMAs are a biostratigraphic system used primarily for the Cenozoic of North America, built upon biostratigraphic relationships among mammals (there are several late Cretaceous NALMAs as well; they are not employed in this evaluation). This system was first established in 1941 by Wood et al.,12 and extensively revised in Woodburne’s mammal compilation.13 It is used extensively in the mammalian paleontological literature, and North American mammal taxa are comprehensively described in relation to the NALMAs in the Woodburne and Janis et al. mammal compilations.8,9,13 Figure 2 shows the relationship of the NALMAs to the stages of the Cenozoic.
One can immediately see from figure 2 that the NALMAs are more numerous than are the stages, and therefore provide finer resolution in biostratigraphic studies than do stages in the Cenozoic. When using the Paleobiology Database for this study, the NALMAs also provide more accurate documentation of both the number of genera reported and the completeness of their respective fossil record, thus providing more accurate biostratigraphic ranges than stages. This is due to the use of NALMAs (rather than stages) for recording mammal occurrences in the compilation texts of Woodburne and Janis et al.,8,9,13 and the uploading of these published data into the Paleobiology Database.
Moreover, the use of the NALMAs also resulted in the removal of non-North American taxa which were occasionally (and curiously) included in stage-based searches of North American mammals. For example, the Giraffidae has no North American taxa (either modern or fossil), yet the biostragraphy of the African members of this group was reported when using stages and limiting the search to North America. Records for giraffes were not encountered when using NALMAs as the search parameter.
Results and analysis
The position of the Pliocene/Pleistocene boundary (dated at 2.6 million years ago by conventional geologists) correlates to the upper portion of the Blancan NALMA (figure 2). For ease of evaluation, the boundary between the Blancan and overlying Irvintonian NALMA will serve as a proxy for the Pliocene/Pleistocene boundary. Genera known from the Irvingtonian NALMA that also record fossils from the Blancan NALMA or below are considered to cross the Flood/ post-Flood boundary when placed at the Pliocene/Pleistocene. Any genus whose highest occurrence is within the Blancan is not considered to cross the boundary. Of the 303 genera surveyed, 70 (23%) cross the Pliocene/Pleistocene boundary. Table 1 summarizes the full analysis, and figures 3 through 11 (figures 4–11, see online supplement14) provide graphical expression for a sample of the families evaluated. In each of these figures, the recovery of a fossil within a genus during one of the NALMAs is represented by a grey square in the centre of the NALMA biozone. The grey square is thus a presence/absence data point during a particular NALMA, and this analysis does not resolve early, middle, or late subdivisions within each NALMA.
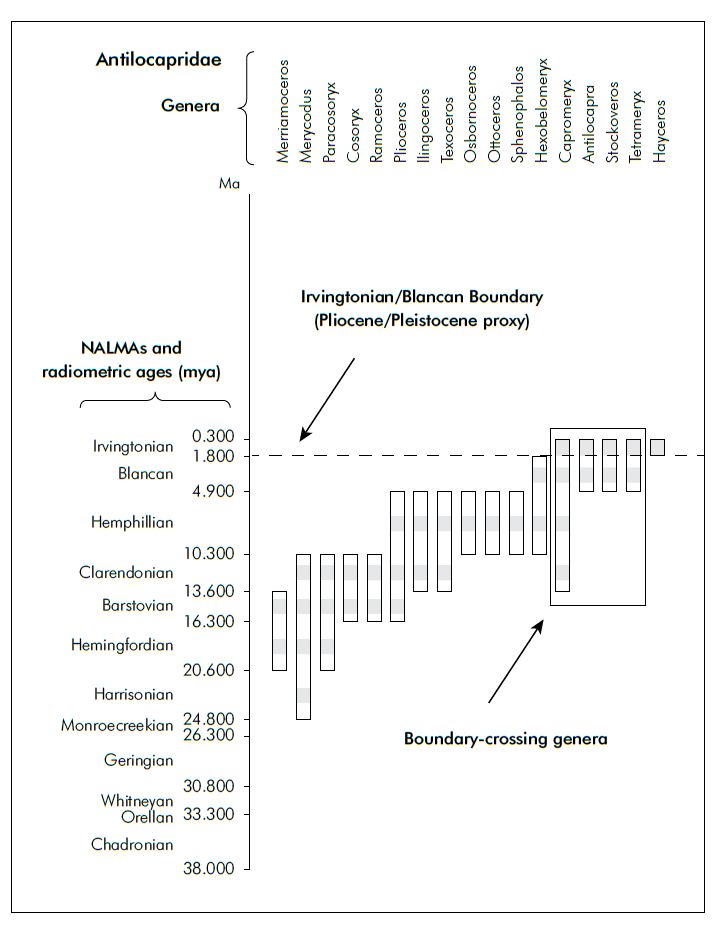
Figure 3 is the output generated for the family Antilocapridae, and serves as a guide for interpreting these figures. Fossils of this unusual ungulate family (which have a bony horn capped by antler material) are found only in North America, and the family is represented today by the lone species Antilocapris americana, the pronghorn antelope. As seen in figure 3, Antilocapris is accompanied by sixteen additional genera from Antilocapridae during the Cenozoic.
A dashed line marks the boundary between the Blancan and Irvingtonian NALMAs, the proxy for the Pliocene/Pleistocene boundary. Of the seventeen total genera, four cross the Pliocene/Pleistocene boundary. Additional examples are provided in figures 4 through 14 (see online supplement14), reflecting a variety of well-known and often-discussed mammalian groups.
When the Pliocene/Pleistocene boundary is used to approximate the Flood/post-Flood boundary, nearly one-fourth of the post-Flood baramin members (understood to be species within the same genus) evaluated here migrated from North America to the Ark, and returned again to North America to coincidentally inhabit the same geographic locations as their pre-Flood (or transported, Flood-buried) baraminic kin.
If pre-Flood baramins are better approximated by the taxonomic rank of family (which is more reflective of current baraminological research and rather broad consensus within the young-earth community), then the situation is far more severe. Twenty-seven of the 28 mammal families studied here include at least one genus which crosses the Flood/post-Flood boundary when placed at the Pliocene/ Pleistocene boundary, and many families display multiple boundary-crossing genera. The lone exception is the Rhinocerotidae (table 1, figure 1214), the last members of which in North America suffer extinction during the Pliocene (figure 10, online supplement14; see also Janis et al.8). So if the family approximates the baramin, then >96% of the mammal baramins evaluated here migrated from Laurentia/North America to the Ark, and returned again to North America.
Moreover, taxa which would have had to return to North America are in some cases genera known only from North America (e.g. Antilocapris (pronghorn antelope), Odocoileus (whitetail and mule deer), Sylvilagus (cotton-tail rabbits)). For these taxa, there is no pool of species from their genus on other continents which could coincidentally migrate to North America during the post-Flood period. In other words: why would endemic pre-Flood North American mammals return only to North America? One would expect, given the ‘sweepstakes’ model of post-Flood migration, that pre-Flood baramins currently known from the Cenozoic of other continents would appear above the boundary as Pleistocene fossils in North America. While it is certainly true that there are a number of genera which appear to migrate from Asia and Europe (especially bovids14) and South America (a number of edentates/xenarthrans), these taxa are themselves known from more recent geological strata, rather than from deeper within the Cenozoic, or below. This supports a Flood/post-Flood boundary significantly lower than the Pliocene/ Pleistocene, with the Cenozoic faunal interchanges and significant endemic development reflective of post-Flood migration and intrabaraminic diversification.
Lastly, and perhaps most damaging: why are there no post-Flood mammal migrants into North America, the Flood-derived fossils of which are otherwise only known from India? Or Africa? Or Australia? The latter is the most damaging case, as the fossil record of mammals in Australia is most unusual, being dominated by an extensive array of marsupials. Yet there are no fossil kangaroos, koalas, or Tasmanian wolves in North America or any other continent. They are not present anywhere else in Pleistocene deposits, or indeed in any other Cenozoic deposits on any of the other continents. A biostratigraphic analysis like this one of the Australian mammal record by a researcher more familiar with these taxa would likely show similar patterns to those seen here, and perhaps even more pronounced.
Conclusions
The biostratigraphic analysis presented here for North American mammalian families makes placement of the Flood/post-Flood boundary at or near the Pliocene/Pleistocene boundary untenable. Rather, these data are more naturally interpreted as representing time-sequential recolonization of the post-Flood world by diversifying terrestrial mammal baramins. Given the biostratigraphic break expected to characterize the Flood/post-Flood boundary, a lower location for the boundary must be sought. At present, the Cretaceous/Paleogene boundary appears to be the stratigraphically highest and most prominent biostratigraphic break (it includes the last inplace stratigraphic appearance of dinosaurs, pterosaurs, and several other bird, mammal, reptile, and amphibian groups), though a similarly thorough analysis must be completed in order to strengthen its claim to the Flood/post-Flood boundary.
| Order | Family | # Genera | # Genera Crossing Plio/Plei Boundary | % Crossing |
|---|---|---|---|---|
| Artiodactlya | Antilocapridae Bovidae Camelidae Cervidae Tayassuidae |
17 16 30 10 11 |
4 1 5 4 2 |
23.5 6.3 16.7 40.0 18.2 |
| Carnivora | Canidae Felidae Hyaenidae Mustelidae Procyonidae Ursidae |
25 15 3 40 7 13 |
3 9 1 7 3 4 |
12.0 60.0 33.3 17.5 42.9 30.8 |
| Edentata/Xenarthra | Dasypodidae Glyptodontidae Megalonychidae Megatheriidae Mylodontidae Nothrotheriidae Pampatheriidae |
2 2 2 1 3 1 1 |
1 2 1 1 1 1 1 |
50.0 100.0 50.0 100.0 33.3 100.0 100.0 |
| Insectovora | Talpidae | 18 | 4 | 22.2 |
| Lagomorpha | Leporidae Ochotonidae |
14 7 |
6 2 |
42.9 28.6 |
| Marsupialia | Didelphidae | 1 | 1 | 100.0 |
| Perissodactlya | Equidae Rhinoceratidae Tapiridae |
27 17 8 |
1 0 1 |
3.7 0.0 12.5 |
| Probocidia | Elephantidae Gomphotheriidae Mammutidae |
1 9 2 |
1 2 1 |
100.0 22.2 50.0 |
| 8 Orders | 28 Families | 303 genera | 70 boundary-crossing genera | 23.1% of genera cross boundary |
Table 1. Genus-level mammalian survivorship across the Pliocene/Pleistocene boundary in North America.
Acknowledgments
I am grateful for the helpful comments and suggestions of the editors and two anonymous reviewers who helped improve this paper. This paper benefitted from numerous engaging discussions with others, especially John Whitmore and Michael Oard at recent Creation Geology Society meetings.
Supplementary Information
References
- Austin, S.A., Baumgardner, J.R., Humphreys, D.R., Snelling, A.A., Vardiman, L. and Wise, K.P., Catastrophic plate tectonics: a global Flood model of Earth history; in: Walsh, R.E. (Ed.), Proceedings of the Third International Conference on Creationism, Technical Symposium Sessions, Creation Science Fellowship, Pittsburgh, pp. 609–621, 1994. Return to text.
- Whitmore, J. and Garner, P., Using suites of criteria to recognize pre-Flood, Flood, and post-Flood strata in the rock record with application to Wyoming (USA); in: Snelling, A.A. (Ed.), Proceedings of the Sixth International Conference on Creationism, Creation Science Fellowship, Pittsburgh, PA, and Institute for Creation Research, Dallas, TX, pp. 425–448, 2008. Return to text.
- Snelling, A., Earth’s Catastrophic Past: Geology, Creation & the Flood, Institute for Creation Research, Dallas, TX, 2009. Return to text.
- Whitcomb, J.C. and Morris, H.M., The Genesis Flood: The Biblical Record and Its Scientific Implications, Presbyterian and Reformed, Phillipsburg, NJ, 1961. Return to text.
- Oard, M., Is the K/T the post-Flood boundary? part 1: introduction and the scale of sedimentary rocks, J. Creation 24(2):95–104, 2010. Return to text.
- Baumgardner, J., Catastrophic plate tectonics: The physics behind the Genesis Flood; in: Walsh R.E. (Ed.), Proceedings of the Fifth International Conference on Creationism, Creation Science Fellowship, Pittsburgh, Pennsylvania, pp. 113–126, 2003. Return to text.
- Oard, M., An Ice Age Caused by the Genesis Flood, Institute for Creation Research, El Cajon, CA, 1990. Return to text.
- Janis, C.M., Scott, K.M. and Jacobs, L.L., Evolution of Tertiary Mammals of North America: Volume 1: Terrestrial Carnivores, Ungulates, and Ungulatelike Mammals, Cambridge University Press, Cambridge, UK, 1998. Return to text.
- Janis, C.M., Gunnell, G.F. and Uhen, M.D., Evolution of Tertiary Mammals of North America: Volume 2: Small Mammals, Xenarthrans, and Marine Mammals, Cambridge University Press, Cambridge, UK, 2008. Return to text.
- Paleobiology Database, www.pbdb.org, accessed 11 April 2011. Return to text.
- For a helpful discussion of the Paleobiology Database, see Karle, K.H., Paleobiology Databases, J. Creation 20(3):87–91, 2006. Return to text.
- Wood, H.E. and Clark, J., Nomenclature and correlation of the North American continental Tertiary, Bulletin of the Geological Society of America 52(1):1–48, 1941. Return to text.
- Woodburne, M.O. (Ed.), Late Cretaceous and Cenozoic Mammals of North America, University of California Press, Berkeley, CA, 1987. Return to text.
- The large number of first appearances in figure 4 (see online supplement at creation.com/pliocene-pleistocene-boundary) reflects significant migration into North America from Asiatic and European bovids. Return to text.
- TSCreator visualization of enhanced Geologic Time Scale 2004 Database (Version 5.0; 2012), James Ogg (database coordinator) and Adam Lugowski (software developer), www.tscreator.org. Return to text.

Readers’ comments
Comments are automatically closed 14 days after publication.