

Journal of Creation 28(1):16–17, April 2014
Browse our latest digital issue Subscribe
More evidence for the reality of genetic entropy
My colleague John Sanford and I have recently published a paper in a secular journal with what we believe are profound implications.1 Our basic claim is that ‘genetic entropy’ works in the real world, which brings questions about the role of natural selection and the long-term survival of species into the future.
A new look at an old virus
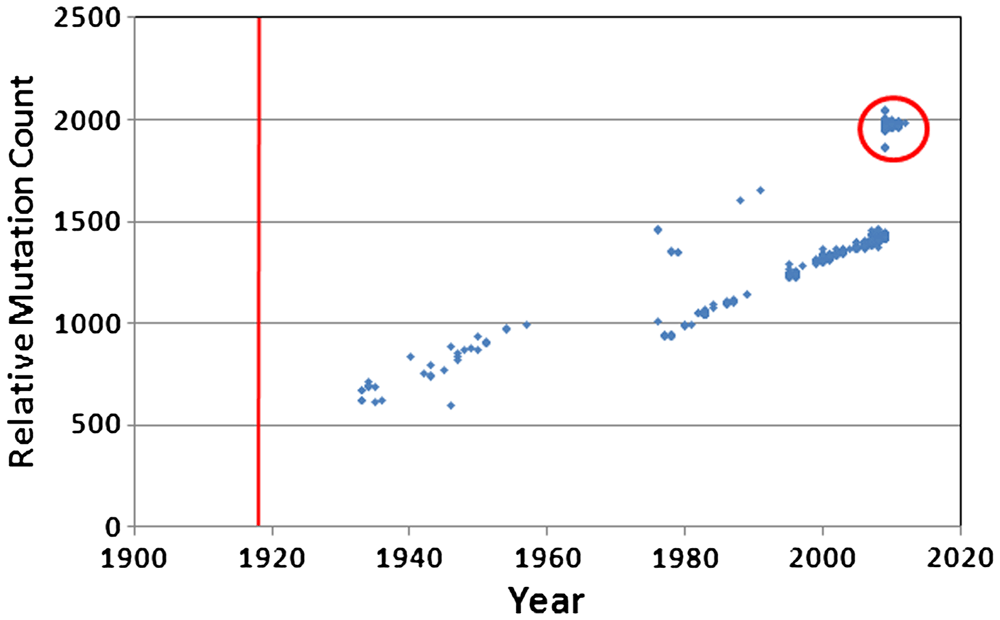
The paper analyzed mutation accumulation in the human H1N1 influenza genome using over 95 years’ worth of genetic sequences (figure 1). This type of data is a rarity in the world of genetics, since most sequence data are from recent organisms with long generation times. The influenza virus, however, has been isolated and sequenced from human tissue samples all the way back to 1918. With a human-to-human transmission on average every three days or so, this makes over 11,000 disease generations and many times more than that number of viral generations. The number of viral generations is possibly comparable to the number of generations since the supposed human-chimpanzee split.

We provided data that more than suggests that the various influenza viruses that infect humans cannot survive long term, and we were the first to notice the disappearance of the human version of the H1N1 influenza virus in mid-2009. We closed our arguments by suggesting government agencies are barking up the wrong tree in their quest to detect newly evolving strains. Instead, scientists should be focusing their efforts to understand the appearance of new viruses, because once a virus makes a cross-species jump it burns fast and hot and eventually burns out. It is the new versions that are the greatest threat, not the old, worn-out ones. We discussed the 2009-2010 ‘swine’ flu outbreak, noting that it was far less serious than expected, and that this was probably due to the fact that the virus (figure 2) had picked up thousands of mutations and was much less robust compared to the original H1N1 ancestral virus that entered the swine population about the same time the human version appeared.
Implications of the study
There are additional implications to our work, although most of these were not explicitly stated in the text.
First, this is apparently the first experiment designed to test the Darwinian mutation/selection model in any species over tens of thousands of generations. All other experiments (even Lenski2) used many fewer generations, or assumed common ancestry (e.g. anything written about the evolution of humans and chimpanzees from a common ancestor) without actually testing it.

Second, despite pervasive and demonstrable natural selection among these viruses, the 1918 version of the human H1N1 virus went extinct, twice, at the appearance of a competing strain, apparently due to a lack of robustness caused by mutation accumulation. The first time was in 1957 when a competing serotype appeared. After an accidental reintroduction of human H1N1 in 1976, the second extinction occurred when a recombined version of the swine H1N1 appeared in humans, and after more than 10% of the human H1N1 genome had ‘rusted away’.
Third, accumulating mutations are not silent, even those that do not affect the amino acid chain of a protein, because codon usage influences translation efficiency. Circulating levels of the many transfer RNAs are proportional to the frequency of their corresponding codons. Thus, changing from common to rare codons is expected to decrease the speed of translation.3 Unchecked mutation accumulation leads to a breakdown in codon bias, potentially affecting translation efficiency in the host cell.
Lastly, since the various mutations accumulated in a linear fashion, those mutations that escaped the selective filter (that would be most of the mutations) apparently accumulated according to the laws of chemistry. Thus, genetic change is overwhelmingly a product of thermodynamics (figure 3), not selection. Over time there was, approximately, a 1% net increase in A, a 0.5% net increase in U (H1N1 is an RNA virus), a 0.5% net loss of C, and a 1% net loss of G.
Natural selection had a minimal effect on mutational direction. Does this mean the direction of ‘evolutionary’ change is predetermined? If so, and if the common ancestor of things like humans and chimpanzees is not ‘on the curve’, they should not have a common ancestor. This would be a very interesting line of study.
Creationists and original research
The reader should note that the editor of the host journal permanently flagged the paper “Highly Accessed” while the provisional paper was still up on their website. Clearly, many in the influenza research community downloaded the paper. So, who says creationists never do original research and who says they never publish in peer-reviewed journals?
References and notes
- Carter, R. and Sanford J.C., A new look at an old virus: patterns of mutation accumulation in the human H1N1 influenza virus since 1918, Theor. BiolMed. Model 9:42, 2012 | doi:10.1186/1742-4682-9-42; tbiomed.com/content/9/1/42. Return to text.
- Blount, Z.D., Borland, C.Z. and Lenski, R.E., Historical contingency and the evolution of a key innovation in an experimental population of Escherichia coli, PNAS 105(23):7899-7906, 2008 | doi:10.1073/pnas.0803151105. See also Batten, D., Bacteria ‘evolving in the lab’?, ‘A poke in the eye for anti-evolutionists’?, 2008; creation.com/citrate. Return to text.
- Xia, X., Maximizing translation efficiency causes codon usage bias, Genetics 144:1309-1320, 1996. Return to text.




Readers’ comments
Comments are automatically closed 14 days after publication.