

Journal of Creation 27(1):7–10, April 2013
Browse our latest digital issue Subscribe
Meiotic recombination—designed for inducing genomic change

Creationary biologists have recognized that the diversity seen within created kinds today cannot be adequately explained by the shuffling of pre-existing gene versions (alleles) and accidental errors that accumulate within the genome.1 Within the context of creation, the development of genetic diversity has been a means by which God has enabled his creatures to adapt to the many different environmental niches they occupy today (Genesis 1:22; 8:17; Isaiah 45:18). Further, it has played an important role in adding variety, beauty, and productivity in various domesticated plants and animals.2
There is certainly no logical reason to believe that unguided chance processes can bring about a functional genome.3 Neither is there sound reason to believe that accidental changes to the genome are a productive source of useful genetic diversity. Logically, therefore, the genome must contain biological information that allows it to induce variation from within.4 One mechanism involved in this is meiotic recombination.5 Continued scientific research is elucidating some amazing details of this process.
Meiosis is a special type of cell division necessary for the formation of gametes (eggs or sperm) so sexual reproduction can take place. In most plants and animals, chromosomes come in pairs (homologs, one derived from each parent), but gametes only carry one of each homolog. Early in meiosis, each chromosome must be drawn to its homolog and stably pair. Then each homolog will be pulled in the opposite direction so that the two cells that form during the division will have exactly one of each homolog.
Meiotic recombination is no accident
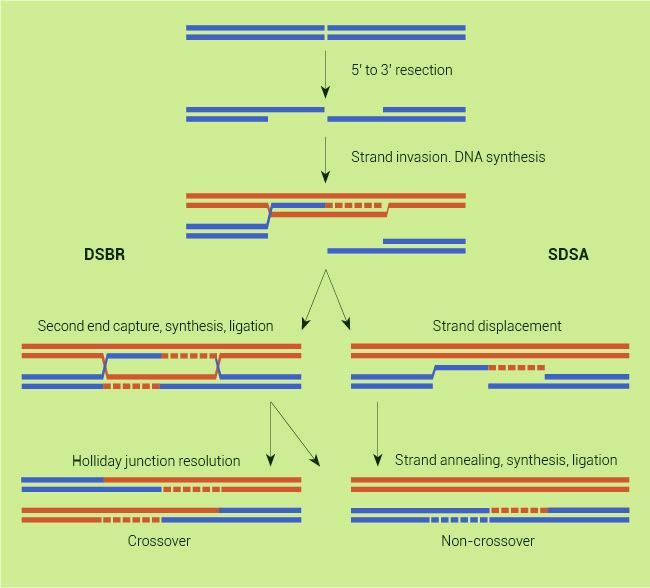
God designed meiosis in a way that naturally tends to increase diversity. In order for the chromosomes to stably pair, recombination occurs between the homologs. The process is initiated by an enzyme which cuts the DNA on one homolog, forming a double-stranded break. Then each side of the break is resected in one direction. This leaves two tails, which are important in repairing the break (figure 1).
There are several pathways by which the break can be repaired. The best known resolution of the break is called crossing over. For this to occur, both of the tails must invade the homolog to form a double Holliday junction (dHJ). DNA synthesis occurs extending these tails. Then, depending on which enzymes are used to cut this structure apart, the distal ends of the chromosomes are swapped.
This swapping between homologs is important in helping to shuffle alleles, which allows for new combinations that may be advantageous. The method of DNA repair described above is known as double-stranded break repair (DSBR). It does not always result in crossing over. A different enzyme can be used to cut the dHJ at a different location and gene conversion will result instead. In gene conversion, a segment from one homolog is copied onto the other. A second pathway for resolving double-stranded breaks is called synthesis-dependent strand annealing (SDSA). In this circumstance only one tail invades the intact homolog and gene conversion is the result.6
Meiotic recombination is mutagenic
Technically, swapping portions of a chromosome and gene conversion are mutations when they alter the nucleotide sequence. Other mutations can also occur during the repair of double-stranded breaks. It appears to be more common with gene conversion. One study in yeast revealed a mutation rate 1,000 times higher during gene conversion than the normal spontaneous mutation rate for that locus. Most mutations were base pair substitutions. About 40% of the mutations were attributable to some form of template switching. In yeast strains with a proofreading defect in a DNA polymerase, template switch mutations were absent.7 This suggests that template switching is a complex, enzyme-driven process.
There is a bias to where meiotic recombination occurs. In a study of Drosophila, crossing over tended to occur in specific hot spots, but these were not influenced by whether or not it was a genic region. Gene conversion had a more uniform distribution, was more common among genic sequences, and was seen where crossing over was rare or absent. The authors emphasized the importance of having information on rates of recombination to include in population genetics models.8 Studies in plants indicate that a variety of genetic and epigenetic factors influence the frequency of crossing over.9
There are several other pathways by which double-stranded breaks can be repaired. One of the most interesting and mutagenic is break-induced replication (BIR). It has been shown to produce complex rearrangements including copy number variation (CNV) and non-reciprocal translocations. These often involve multiple rounds of template switching. Specific endonucleases are necessary for proper BIR; an absence of these endonucleases has been shown to significantly reduce template switching.10
Significance of mutations
At times mutations are explained as the result of accidents which introduce errors into the DNA sequence. The concept of non-directed change is foundational in the standard evolutionary model. Logically, accidental changes in a complex system should be consistently harmful to some degree. Creationists have pointed this out in emphasizing the implausibility of accidents in accounting for the complexity of life.
However, when diversity is examined within a creation model, it is evident that significant diversity has arisen since the time of the Flood. In contrast to the notion that all mutations are harmful, the observed diversity does not appear to be typically harmful, and much is considered to be healthy. It has been pointed out that this useful diversity is not logically the result of accidents, but some designed mechanism(s) must be producing it.1
Several specific examples are worth noting. In a gene influencing coat colour, a pattern of in-frame indel (insertion or deletion) mutations was noted across several unrelated kinds. These generally result in a black coat colour. Statistically, only one in three indels should be in-frame. It does not appear that natural selection can explain this bias toward in-frame indels, and so a designed mechanism was suggested as its source.11
Resistance to organophosporous insecticides has been studied in sheep blowflies. There is a particular gene where specific mutations can confer resistance to one organophosphate or another. Resistance to one of these insecticides (malathion) was identified in pinned specimens that pre-dated the first use of that insecticide; therefore, selection would be a reasonable explanation for how it spread in the fly population. Resistance to a second insecticide (diazinon) appears to have arisen by mutation since the insecticide was introduced. This rapid appearance of resistance is quite impressive (though disheartening for those trying to get rid of this pest). In addition to this, flies have emerged that are resistant to both insecticides as a result of gene duplication (a form of CNV). It appears that such gene duplications have arisen at least three separate times in these flies, and always involve the resistant alleles.12
The point here is that the mutagenic nature of meiosis appears to provide a plausible mechanism for inducing this type of variation within a creationary timeframe. The requirement of specific enzymes and non-random pattern of change in meiotic recombination suggests it could play a significant role in producing the observed useful genetic diversity.
Gene conversion, a designed mechanism which can result in fixation of alleles
Gene conversion can lead to a transmission distortion, a deviation from the expected ratio of alleles in the gametes. Studies in mice revealed an example of this due to a preferential induction of double-stranded breaks on one homolog, which yielded an over-transmission of the allele from the other. Given the distortion, population simulations predicted that the favoured allele would be fixed in the population in less than 1,200 generations.6
Transmission distortion is extremely significant. Most models attempting to explain the changes in allele frequency of a population assume that a heterozygous parent would have an equal chance of passing either of the alleles on to the offspring. The fixation of alleles within a population is generally attributed to natural selection, although genetic drift is also recognized as a possibility. These are naturalistic explanations that fit well within the ‘anti-designer’ presuppositions of the evolutionary model.
Despite the appeal of scenarios crediting natural selection, they may have little semblance to reality if designed mechanisms are involved in changing allele frequencies. One example in animals would be migration. Perhaps animals move to where they are most comfortable because God gave them the wisdom to do so, thus enabling them to survive and reproduce. This comfort factor may be related to having a genotype compatible with (adapted to) that environment. So essentially, animals with adaptive alleles stay, and the others leave. This is rather the reverse of natural selection (where the environment ‘selects’ the animals), as it is the animal making a conscious choice.
Transmission distortion due to gene conversion, as described above in mice, may also prove to be an important mechanism for fixation of adaptive alleles in populations. If this turns out to be the case, it is a serious problem for evolutionists. It would be another major blow to the view that naturalistic processes adequately explain the origin of new species. Instead, designed mechanisms would be important for both the generation of diversity and fixing adaptive alleles within a population. If designed processes are necessary for adaptive changes even within created kinds, it points again to an awesome Creator!
Summary
One thing is clear; the evolutionary based inference that mutations (any change in the DNA sequence) are always accidents or copying errors is false. Changes in DNA sequence can arise for a number of reasons. One reason is that meiotic recombination, an essential step in reproduction for many plants and animals, is designed to induce genetic changes. This is highlighted by the fact that enzymes are necessary for this complex processes, including enzymes which induce the double-stranded breaks and facilitate template switching. Since this is the case, I fully expect that better understanding meiotic recombination will be one piece in the puzzle to better understanding how diversity has risen so quickly within created kinds since the time of the Flood.
References
- Lightner, J.K., Karyotypic and allelic diversity within the canid baramin (Canidae), J. Creation 23(1):94–98, 2009. Return to text.
- Lightner, J.K., The effect of mutations down on the Farm, Answers in Depth 5(1), 2010, www.answersingenesis.org/articles/aid/v5/n1/effect-of-mutations-down-on-farm, accessed 27 December 2012. Return to text.
- Such reasoning only ‘makes sense’ if one is grossly ignorant of the complexity of the genome and/or refusing to consider the possibility of a creator; Romans 1:16–22. Return to text.
- Terborg, P., The design of life: part 3 an introduction to variation-inducing genetic elements, J. Creation 23(1):99–106, 2009. Return to text.
- Ashcraft, C.W., Genetic variability by design, J. Creation (formerly TJ) 18(2):98–104, 2004. Return to text.
- Cole, F., Keeney, S. and Jasin, M., Preaching about the converted: how meiotic gene conversion influences genomic diversity, Annals of the New York Academy of Sciences 1267:95–102, 2012. Return to text.
- Malkova, A. and Haber, J.E., Mutations arise during repair of chromosome breaks, Annual Review of Genetics 46:455–473, 2012. Return to text.
- Comeron, J.M., Ratnappan, R. and Bailin, S., The many landscapes of recombination in Drosophila melanogaster, PLoS Genetics 8(10):e1002905, 2012. Return to text.
- Henderson, I.R., Control of meiotic recombination frequency in plant genomes, Current Opinion in Plant Biology 15:556–561, 2012. Return to text.
- Pardo, B. and Aguilera, A., Complex chromosomal rearrangements mediated by breakinduced replication involve structure-selective endonucleases, PLoS Genetics 8(9):e1002979, 2012. Return to text.
- Lightner, J.K., Genetics of coat color I: The melanocortin 1 receptor (MC1R), Answers Research J. 1:109–116, 2008. Return to text.
- Lightner, J.K., Pattern of change over time: organophosphorous resistance in the Australian sheep blowfly, Lucilia cuprina, J. Creation 22(1):81–84, 2008. Return to text.




Readers’ comments
Comments are automatically closed 14 days after publication.