
The Hobbit: Precious fossil or poisoned chalice?
22 June 2006
When Homo floresiensis was announced as a new ‘hominid’ species in the prestigious journal Nature at the end of October 2004, it must have been a euphoric moment for the discoverers.1 At the time the authors believed they were dealing with a dwarfed Homo erectus species, and because of the small size of the main fossil specimen (LB1) the species was dubbed ‘the Hobbit.’ Birthday suit pictures of this supposed ape–man, after a successful rat hunt, were splattered around the world, as newspapers and other media outlets announced that a lost race of human ‘hobbits’ had been unearthed on an Indonesian island.2
The discoverers were suddenly the stars of paleoanthropology, as their little Hobbit took to the imagination of people the world over. Reconstructed using considerable artistic license and imagination, the Hobbit even made the cover of February 2005 Scientific American and April 2005 National Geographic. If only it could have ended on this romantic note, but alas—as the fanfare subsides, the Hobbit looks decreasingly like a diminutive character in JRR Tolkien’s novels, and increasingly like a case study in human pathology.
Why not an australopithecine?
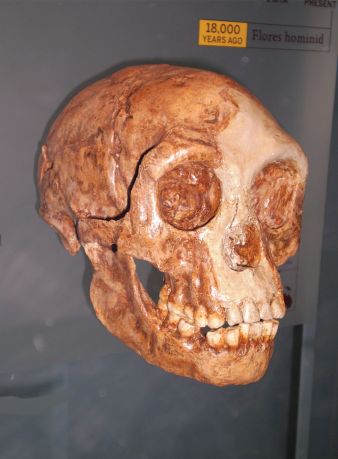
At a tiny stature of just over one meter high, and with a very small brain capacity of about 400 cm3, one would perhaps have expected evolutionists to push for the inclusion of the Hobbit as a member of the australopithecines. Why didn’t they? Firstly, its supposed age of 18,000 years,3 and location in Flores, Indonesia, are from an evolutionist standpoint temporally and spatially outside the known australopithecine range respectively.4 Evolutionists currently believe that the last of the australopithecines died out over a million years ago, and so if the Hobbit was an australopithecine it would almost be like finding a ‘living fossil.’ A bigger problem is that the australopithecines are believed by evolutionists to have originated in Africa, and so for them get to Flores, Indonesia, means they would have had to build watercraft, a feat seemingly beyond their brainpower. However, there are also anatomical reasons for believing they were not australopithecines.
The discoverers describe the body proportions of LB1 as being the same as AL288–1 (Lucy) Australopithecus afarensis, but unlike all other supposed hominids (including Homo erectus) ‘for which there are reliable data.’5 In the original Nature paper the ilium of the LB1 pelvis was described as having a ‘marked lateral flare’ compared with modern humans.6 This is more a characteristic of the australopithecines, as deduced from the reconstructed pelvis of the afarensis specimen known as ‘Lucy.’7 In their follow–up paper, about a year later, the pelvis is described as ‘flared antero–laterally, consistent with an australopithecine–shaped thoracic region.’5 However, a more recent examination and reconstruction of the LB1 pelvis by William Jungers of Stony Brook found ‘the orientation of the pelvic blades’ to be similar to ‘modern’ humans.8 According to Indonesian paleoanthropologist Teuku Jacob of Gadjah Mada University in Jakarta, who examined some of the LB1 skeletal remains under controversial circumstances,9,10,11 the pelvis and the thigh bone were human, with the only abnormalities being in the skull.12 He found that the ‘legs, arms and everything else were genetically normal.’12
Regarding similarity, it is stated about LB1 that in ‘the overall shape of its skull and its teeth, the creature most closely resembles Homo erectus.’13 According to the authors of the floresiensis paper LB1 ‘does not have the great postcanine tooth size, deep and prognathic facial skeleton, and masticatory adaptations common to members’ of the genus Australopithecus.14 They argue that:
‘Instead, the facial and dental proportions, postcranial anatomy consistent with human–like obligate bipedalism, and a masticatory apparatus most similar in relative size and function to modern humans all support assignment to the genus Homo—as does the inferred phylogenetic history, which includes endemic dwarfing of H. erectus.’15
Compared to modern humans the arms of LB1 are long with respect to the legs.16 The limb proportions, as indicated by the humerofemoral index (ratio of humerus to femur length), have been measured to be the same as the AL288–1 afarensis skeleton, and outside the range of variation normally attributed to humans.5 The estimated humerofemoral index in afarensis, although less than that of extant apes, is still significantly greater than that of humans.17 Hence, the limb proportions of LB1 are australopithecine–like. However, according to Leroi, ‘Careful measurements of pygmies (and thousands of them have been measured) show that compared to taller people, pygmies have relatively short legs but relatively long arms.’18 Hence, it may be that as the human body is ‘sized’ down the limbs do not scale down proportionally.
Interestingly, Carl Vogt, a prominent anthropologist and early evolutionist of the late Victorian era, who studied microcephaly (a pathological condition in which the brain fails to develop to its normal size) thinking it would help him understand supposed evolution in humans, commented in his Lectures On Man that in certain microcephalics the ‘arms seem disproportionately long, the legs short and weak.’19
According to anthropologist Alan Thorne, Australian National University in Canberra, dwarfism ‘goes with microcephaly, especially in hunter/gatherer populations.’12 Dwarfism commonly results from a deficiency of growth hormone secretion from the anterior lobe of the pituitary gland in the brain. In this condition:
‘In general, all the physical parts of the body develop in appropriate proportion to one another, but the rate of development is greatly decreased. A child who has reached the age of 10 years may have the bodily development of a child aged 4 to 5 years, and the same person at age 20 years may have the bodily development of a child aged 7 to 10 years.’20
An example of someone who most likely had this condition was Joseph Boruwlaski (1739–1837). Portraits of this able person indicate he was ‘perfectly proportioned in his smallness.’21 He continued growing slowly until he was thirty, after which his height remained fixed at 3 feet 3 inches (99 cm).22 For comparison, the LB1 skeleton had a height estimated as 106 cm.14 Dwarfism and microcephaly occurring in the same individual may not be that unlikely, as severe microcephaly is a condition with a very small head and an undeveloped brain, so it is possible that the pituitary gland in the brain becomes dysfunctional because of the brain pathology associated with severe microcephaly.
If a microcephalic skull was placed on a postcranial skeleton like that of Boruwlaski, and the remains buried in the hobbit ‘pit’ in Flores at the same time as LB1, an interesting question would be whether the same discoverers would be claiming another hobbit. Charles Stratton (1838–1883), at 40 inches tall (101.5 cm), was another mentally capable person of small stature who was ‘well–proportioned throughout his body.’23 According to Barnhart a ‘profile view of his head indicates facial characteristics with many of the same allometric differences seen in LB1.’24
What about microcephaly?
A few days after the hobbit announcement, paleopathologist25 Maciej Henneberg of the University of Adelaide claimed in a newspaper article that the LB1 individual suffered from a pathological growth condition called secondary microcephaly, and that ‘the skull of the Flores hominid is very similar to a 4,000–year–old microcephalic modern human skull found on the island of Crete.’ Henneberg, along with Alan Thorne, soon outlined their case in the journal Before Farming.26 In the same issue, two of the authors of the Nature ‘hobbit’ paper, Peter Brown and Mike Morwood, denounced the ‘research’ by Henneberg and Thorne as ‘extremely poorly informed, and ill designed.’27 Not to be outdone, Thorne later commented that if ‘it’s a case of microcephaly, there are a lot of people in my field who cannot recognise a village idiot when they see one.’12
Also, Teuku Jacob joined the hobbit ‘bashers’ by claiming ‘the specimen was a diminutive modern human.’16 More specifically, Jacob also believes LB1 suffered from microcephaly, and as an explanation for the small size of the skeleton points out that there is evidence of pygmy peoples in the region.12 Jacob led an anthropology research team there and found a pygmy community in the village of Rampapasa, about 1 km from the village of Liang Bua, where the Hobbit was found.28 In this community’ 80 per cent of the Rampapasa villagers were small, with most male adults under 145 cm and female adults about 135 cm.’28 Hence, it is possible that LB1 was just a very small member of such a pygmy population, but additionally suffered from microcephaly. According to paleoanthropologist John Hawks, Jacob’s conclusion that LB1 is a microcephalic pygmy ‘is supported by the observation that pygmies are common in the region today, may have been common in the past, and the possibly high frequency of secondary microcephaly, induced by nutritional deficiency or other non–genetic factors.’29
To attempt to silence some of the critics, the brain of LB1 was assessed by comparing its virtual endocast with endocasts from Homo sapiens, Homo erectus, a human microcephalic, a human pygmy, great apes, Australopithecus africanus (Sts 5), and Australopithecusaethiopicus30 (WT 17000).31 This study, led by anthropologist Dean Falk, concluded that LB1’s ‘well–convoluted brain’ was not a microcephalic or pygmy, but in shape resembled erectus, and additionally they estimated a 417 cm3 virtual cranial capacity for LB1.32 But there was only one microcephalic sample, a plaster–based cast of a skull traced back to the original skull of a 10–year–old boy (Jakob Moegele) with a cranial capacity of 272 cm3.33 Hence, from such a limited sample (one), it seemed premature to claim that LB1’s brain and skull was the wrong shape to be a microcephalic.34
About six months later a brief paper was published by Weber et al, detailing the analysis of 19 microcephalic modern humans, where the finding of a microcephalic endocast comparable to LB1 was reported.35 The brain volume of the microcephalics varied between 290 and 591 cm3, with a mean of 404 cm3. Based on their study, the authors stated that:
‘Both skull and brain morphologies of microcephalics are extremely heterogeneous [varied, diverse] and grossly resemble the anatomy and proportions of H. floresiensis.’35
‘Widely differing index measurements’ were observed in the microcephalic brains, with the indices of one of them resembling aethiopicus.35 Unsurprisingly, the research team led by Falk disagreed with the above assessment by Weber et al, citing lack of information about measurement indices as one of the main issues.36 More recently, Martin et al entered the hobbit fray when they argued against LB1 being a dwarf derived from erectus, as they calculated LB1’s predicted body size when dwarfed, based on its small cranial capacity, to be much smaller than it actually was.37 Instead, they also suggested that LB1 was a microcephalic modern human and cited two microcephalic skulls and endocasts (Indian Hunterian male and Lesotho woman specimens) said to be similar to LB1. As expected, Falk’s group disagreed with the Martin et al assessment, this time citing lack of detail on line drawings of the endocasts as one of the main reasons.38
One thing to come out of the Weber et al study is that any notion that microcephalics have typically simplified gyral patterns or a certain brain shape is wrong, as the authors reported great variability in overall microcephalic brain shape and convolution patterns.35
The finding of fossils from non–microcephalic intelligent hobbit–size people with a cranial capacity of about 400 cm3 would question the notion of an arbitrary cerebral rubicon in the 600–800 cm3 brain size range39 that must be passed in order to have a human mental faculty. The average brain size of chimpanzees is 383 cm3, orangutans 404 cm3, and gorillas 504 cm3.40 Hence, the 400 cm3 brain size of floresiensis from the Indonesian island of Flores is very small if it is non–pathological, yet it possesses human–like intelligence, as adduced by the tools. However, when considering brain size one should also take into account body size. This is done by calculating a value known as the encephalization quotient (EQ).41 According to the discoverers, if the body of floresiensis specimen LB1 is assumed to be lean and narrow, then the estimated EQ places LB1 easily within the erectus range.14
Do multiple hobbits disprove the microcephaly hypothesis?
Although a second skull has yet to be found, a second adult mandible has been discovered, as has postcranial material from other individuals.42 The researchers think that their sample of hobbit bones is from at least nine individuals, with the new mandible described as ‘extraordinarily similar to the first one.’43 On the surface this would seem to rule out the microcephaly theory, but some types of microcephaly may run in families. For example, three of the ten siblings of the microcephalic boy Jakob Moegele were also microcephalics.37 Another example is the tragic case of the three microcephalic brothers used as ‘hominid ancestor’ throwback exhibits at an amusement park in Bangalore, India.44 In the paper by Martin et al the authors state:
‘In addition to genetic factors increasing the likelihood of microcephalics occurring together, it is conceivable that cultural factors might have enhanced this, as at a recent religious site to which microcephalics were brought.’37
The finding of another lower jaw (LB6/1),45 estimated to be 15,000 years old,46 as opposed to the 18,000 years old date for the LB1 skeleton (including its lower jaw), would make the argument of running in the family, if it was for thousands of years, difficult to swallow. However, this assumes that the datings are correct. Evolutionist Robert Martin, of The Field Museum, Chicago, and advocate of the microcephalic hypothesis, in response to two floresiensis lower jaws having been unearthed and supposedly dated thousands of years apart, questions the degree of similarity of the jaws as well as ‘whether the dating of the remains is accurate.’47 Martin has every right to question the accuracy of the dating—a privilege not usually extended to non–believers in evolution, of course.
If the Hobbit turns out to be a microcephalic modern human then one may legitimately ask what other supposed ‘hominids’ are simply microcephalic humans— particularly some of the smaller Homo erectus/ Homo habilis crania. According to its discoverers, the LB1 skull is most reminiscent of smaller erectus fossils, such as ‘the 1.77–million–year–old Dmanisi people from Georgia, in Western Asia’, with both the Dmanisi people and hobbit having skulls that are ‘pinched in at the temples.’48 Hence, it is conceivable that the Dmanisi finds, including four skulls,49 also represent microcephalics occurring together, whether for family, religious or cultural reasons. An ‘enormous’ lower jaw (D2600), unearthed at Dmanisi in 2000, and described as ‘far too large to fit comfortably with any of the crania yet discovered’,50 indicates that people of ‘normal’ brain size (non–microcephalic) also inhabited the region at the same time.
It should be emphasized that just because people are microcephalic does not necessarily imply that they are retarded mentally. For example, evolutionists Skoyles and Sagan refer to a woman with an estimated brain volume of 760 cm3 who had an IQ of 112, which is certainly not retarded.51 Also, there is enormous variation in skull size and shape in so–called ‘modern humans’, and there likely was even greater variation in the past. Add environmental and dietary factors, and many of the alleged ‘hominids’ in the genus Homo, such as Neandertals, erectus and other so–called ‘archaics’, can be explained as simply variations within the human population that descended from Adam and Eve. Hence, it is possible that the hobbit and Dmanisi people were microcephalic ‘erectus’ type humans, but fully human nonetheless.
Susan Anton of New York University believes there is little difference between LB1 and erectus, at least not enough to warrant a new species, as ‘the difference in shape between LB1 and Homo erectus is less striking than that between a Great Dane and a Chihuahua.’52 She is then quoted by Wong as observing that:
‘The possibility exists that the LB1 specimen is a H. erectus individual with a pathological growth condition stemming from microcephaly or nutritional deprivation.’53
Concerning the remains of other individuals at Liang Bua, the estimated sizes of these individuals vary greatly. For example, Henneberg and Thorne estimated that a radius from the site corresponded to a stature of 151–162 cm, depending on how it was reconstructed, which ‘is by no means a dwarfed stature.’54 On the other hand, Jungers notes that limb bones from other individuals ‘are even smaller—“they make LB1 look like the Hulk,” he says.’8
The toolmaker of Flores
In the same issue of Nature that first described the LB1 floresiensis fossil, a companion article included details of dating methods and tools associated with the fossil site in Liang Bua, on the island of Flores, Indonesia.55 Stone artifacts were reported as occurring at the same stratigraphic levels as the LB1 skeleton, including in the same 2 by 2 metre sector where the skeleton was found.56 In the words of Kate Wong, ‘a suite of sophisticated artifacts’ was found at the same level as the skeleton, ‘including awls, blades and points—exhibiting a level of complexity previously thought to be the sole purview of H. sapiens.’ 52 Additionally, significant brainpower would be required to build the boats or watercraft needed to colonize Flores, which up to then evolutionists believed only humans were capable off.57 Evidence indicates that the tool makers at Liang Bua were also able to use fire and hunt stegodonts,58 indicating they were very intelligent. If the hobbit was severely microcephalic and hence retarded, which now seems likely, the tools probably belonged to non–microcephalic humans of the same human population.
The finding of stone tools on Flores has previously been reported (at Mata Menge), but they were age dated to between 0.88 and 0.80 million years old, and on the basis of these ages the artefacts were attributed to being ‘produced by Homo erectus rather than Homo sapiens.’59 Such jumping to conclusions is striking, but being locked into the evolutionary viewpoint gives the authors little alternative.
Following more recent excavations in Mata Menge a paper was published claiming similarity in the stone artefacts from the alleged 0.88 and 0.80 million years old deposits at Mata Menge and those found in the supposed 95 to 12 thousand years old deposits at Liang Bua cave (associated with floresiensis).60 Once again, although no ‘hominid’ remains have been recovered at the Mata Menge site, the authors concluded that ‘the age of the site clearly precludes modern humans.’61 They suggested that the ‘most parsimonious explanation’ was that the stone artefacts from both sites (Mata Menge and Liang Bua) ‘represent a continuous technology made by the same hominin62 lineage.’ 61 Rather than admit the manifestly obvious and most logical conclusion, that humans had to be responsible for both sets of stone artefacts (and also that the huge alleged gap in age between the two tool sites is doubtful), and thus negating the idea of an evolutionary lineage of hominids, the authors state that the ‘Mata Menge evidence negates claims that stone artefacts associated with H. floresiensis are so complex that they must have been made by modern humans (Homo sapiens).’63
Such bewildering logic is perhaps best summed up by evolutionists John Gribbin and Jeremy Cherfas when, under different circumstances, they commented that ‘we must admit that the history of palaeontology does not read as a shining example of the pursuit of truth, especially where it was the truth of man’s origins that was at issue.’64 Rather, it is a search for a purely naturalistic explanation, regardless of whether this is plausible. As indicated by evolutionist John Reader: ‘Preconceived notions have played a fundamental role in the study of fossil man.’65 The above example is a good illustration of how the interpretation of artefacts and fossils is heavily biased according to the researchers’ framework. The reality is that scientists, both evolutionists and creationists, are biased when it comes to studying the origin of man and life, as they interpret the evidence through their own peculiar worldview or framework. The problem arises when the researcher erroneously believes their worldview is ‘science’, and hence objective, whilst alternative viewpoints are ‘religious’, and therefore biased.
What else could the Hobbit be?
Rather than a dwarfed Homo erectus, some researchers have proposed possible alternative ancestors to floresiensis, including Homo habilis,66 or even an ‘offshoot of Australopithecus’, as suggested by Milford Wolpoff.4 However, such notions are purely speculative, being firmly based on the assumption of evolution, not on any evidence.
The team who unearthed the find originally suggested that floresiensis may have been the descendants of erectus from the nearby island of Java, believed to have been there as long as 1.6 mya.67 They suggested that the first hominid immigrants to Flores ‘may have had a similar body size to H. erectus and early Homo, with subsequent dwarfing; or, an unknown small–bodied and small–brained hominin may have arrived on Flores from the Sunda Shelf.’ 14 However, even these researchers now believe erectus was too big to evolve into the diminutive Hobbit.68 For example:
‘Michael Morwood of the University of New England in Armidale, Australia, says he believes the hobbits may instead descend from a smaller, as–yet–undiscovered hominid, resembling 1.8 million–year–old specimens found at Dmanisi in Georgia.’68
The principal author of the first hobbit article, Peter Brown of the University of New England, is quoted as leaning towards the idea that ‘there was some more australopithecine–like ancestor involved.’69 ‘Progressive creationists’, such as Fazale Rana and Hugh Ross, appear to accept that ‘H. floresiensis, like the Neanderthals, coexisted with modern humans.’70 However, the only thing certain about their human origins concept (spirit–less quasi–humans created prior to Adam and Eve) seems to be that no finding causes any difficulty for it, as illustrated by the following ‘hand waving’ statement:
‘This unusual and unexpected discovery causes little difficulty for the RTB human origins model. H. floresiensis is clearly distinct from modern humans, not only in morphology, but in behavior as well. Like H. erectus, H. floresiensis behaved in nonhuman ways. The RTB model considers these hominids in the same vein as the great apes—nonhuman creatures made by God (before He created human beings) that later became extinct.’70
Despite their small size, the remains attributed to floresiensis were initially suggested by creationist Carl Wieland as likely to be descendants of Adam nonetheless.71 In this alternative view floresiensisis ‘a miniature human being exhibiting part of the same range of post–Babel human variation as encompasses the larger so–called Homo erectus.’72 This was also the argument of creationist Kurt Wise,73 and has yet to be ruled out, but in both instances this view was based on the notion that the Flores skeleton was non–pathological. As arguments to the contrary accumulate, it is probably more likely that the LB1 skeleton, whilst still human and hence a descendant of Adam, is a reflection of post–Babel human pathological variation. One thing seems certain; we have not heard the last of this hobbit tale.
Editor’s note: For earlier articles on this site about the ‘hobbit’, see:
About the author
Dr Peter Line’s undergraduate major was in biophysics. After that he completed a masters degree and a Ph.D., both in the area of neuroscience. He has had a keen interest in the creation/evolution issue ever since becoming a Christian, as evolution was a stumbling block to him believing God’s Word was true. Return to top.References and notes
- Brown, P., Sutikna, T., Morwood, M.J., Soejono, R.P., Jatmiko, E. W. Saptomo and Due, R.A., A new small–bodied hominin from the late Pleistocene of Flores, Indonesia, Nature 431:1055–1061, 2004. Return to text.
- See, for example: Cauchi, S., Lost race of human ‘hobbits’ unearthed on Indonesian island, The Age, p. 1, Thursday 28 October, 2004. Return to text.
- Morwood, M.J. et al., Archaeology and age of a new hominin from Flores in eastern Indonesia, Nature 431:1088, 2004. Return to text.
- Wong, K., The littlest human, Scientific American 292(2):46, 2005. Return to text.
- Morwood M.J. et al., Further evidence for small–bodied hominins from the Late Pleistocene of Flores, Indonesia, Nature 437:1016, 2005. Return to text.
- Brown et al., ref. 1, p. 1058. Return to text.
- Mehlert, A.W., Australopithecines – the extinct southern apes of Africa: a fresh light on their status?, J. Creation 14(3):94, 2000. Return to text.
- Culotta, E., How the hobbit shrugged: Tiny hominid’s story takes new turn, Science 312:984, 2006. Return to text.
- Culotta, E., Battle erupts over the ‘Hobbit’ bones, Science 307:1179, 2005. Return to text.
- Culotta, E., Discoverers charge damage to ‘Hobbit’ specimens, Science 307:1848, 2005. Return to text.
- Dalton, R., Fossil finders in tug of war over analysis of hobbit bones, Nature 434:5, 2005. Return to text.
- Vidal, J., Bones of contention, The Guardian, 13 January 2005, guardian.co.uk, 21 January 2005. Return to text.
- Knight, W. and Nowak, R., Meet our new human relatives, New Scientist 184(2471):8, 2004. Return to text.
- Brown et al., ref. 1, p. 1060. Return to text.
- Brown et al., ref. 1, pp. 1060–1061. Return to text.
- Balter, M., Skeptics question whether Flores hominid is a new species, Science 306:1116, 2004. Return to text.
- Richmond, B.G., Aiello, L.C., Wood, B.A., Early hominin limb proportions, J. Human Evolution 43:534–535, 2002. Return to text.
- Leroi, A.M., Mutants: On the form, varieties and errors of the human body, Harper Perennial, London, p. 186, 2005. Return to text.
- Vogt, C., Lectures On Man: His Place in Creation, and in the History of the Earth, Longman, Green, Longman, and Roberts, Paternoster Row, London, pp. 195, 1864; Vogt actually was referring to the microcephalics as idiots. An idiot is a person of profound mental retardation. However, the term belongs to a classification system of an earlier era, and is now considered offensive. Idiocy may result from an undeveloped brain, which is termed microcephalic. It is in the context of microcephaly that Vogt appears to be using the term, as the idiots are described as having a ‘forehead nearly absent’ (p. 195) and a head ’disproportionately small in comparison with the body’ (p. 196). Return to text.
- Guyton, A.C. and Hall, J.E., Textbook of Medical Physiology, eleventh edition, Elsevier Saunders, Philadelphia, Pennsylvania, p. 927, 2006. Return to text.
- Leroi, ref. 18, pp. 172, 175. Return to text.
- Leroi, ref. 18, pp. 174. Return to text.
- Barnhart, W.R., Tiny skeletons and human evolution, Creation Matters 11(1):1–3. Return to text.
- Barnhart, ref. 23, p. 3. Return to text.
- A paleopathologist diagnoses diseases in the past based on the evidence of fossil bones. Return to text.
- Henneberg, M. and Thorne, A. Homo floresiensis: reactions and replies, Before Farming (online journal) 4, article 1, pp. 2–4, 2004. Return to text.
- Brown, P. and Morwood, M. Homo floresiensis: reactions and replies, Before Farming (online journal) 4, article 1, p. 6, 2004. Return to text.
- Pygmy found near home of hobbits, heraldsun.com.au, 6 May 2005.
- Hawks, J., The Liang Bua debate, continued, 16 January 2005, johnhawks.net, accessed January 2005. Return to text.
- The authors of the paper have named this specimen (also known as the ‘Black Skull’) as Paranthropus aethiopicus, but others consider it as part of the genus Australopithecus. Return to text.
- Falk, D. et al. The Brain of LB1, Homo floresiensis, Science 308:242–245, 2005. Return to text.
- Falk et al., ref .31, p. 242, 245. Return to text.
- Martin et al., Comment on “The Brain of LB1, Homo floresiensis”, Science 312:999b, 2006. Return to text.
- Balter, M., Small but smart? Flores hominid shows signs of advanced brain, Science 307:1387, 2005. Return to text.
- Weber, J., Czarnetzki, A. and Pusch, C.M., Comment on “The Brain of LB1, Homo floresiensis”, Science 310:236b, 2005. Return to text.
- Falk, D., et al., Response to Comment on “The Brain of LB1, Homo floresiensis”, Science 310:236c, 2005. Return to text.
- Martin et al., Comment on “The Brain of LB1, Homo floresiensis”, Science 312:999b, 2006. Return to text.
- Falk, D. et al., Response to Comment on “The Brain of LB1, Homo floresiensis”, Science 312:999c, 2006. Return to text.
- Leakey, L.S.B., Tobias, P.V. and Napier, J.R., A new species of the genus Homo from Olduvai Gorge, Nature 202:7, 1964. Return to text.
- Tobias, P.V., Evolution of the human brain; in: Gordon, E. (Ed), Integrative Neuroscience, Harwood Academic Publishers, Amsterdam, p. 39, 2000. Return to text.
- Aiello, L. and Dean, C., An Introduction to Human Evolutionary Anatomy, Academic Press, London, pp. 189–191, 1990. Return to text.
- Morwood et al., ref. 5, pp. 1012–1017. Return to text.
- Lieberman, D.E., Further fossil finds from Flores, Nature 437:957, 2005. Return to text.
- Lesko, J.P., Fun World amusement?, J. Creation 17(3):58, 2003. Return to text.
- Morwood et al., ref. 5, pp. 1014. Return to text.
- Lieberman, ref. 43, p. 958. Return to text.
- Wong, K., Salvos exchanged in battle over hobbit brain, 25 May 2006, blog.sciam.com, accessed 2 June 2006. Return to text.
- Morwood, M.J., Sutikna, T. and Roberts, R., The people time forgot, National Geographic 207(4):7, 12, April 2005. Return to text.
- Fischman, J., Family Ties, National Geographic 207(4):16–27, 12, April 2005. Return to text.
- Wong, K., Stranger in a New Land, Scientific American 289(5):56, 2003. Return to text.
- Skoyles, J.R. and Sagan, D., Up from dragons: The evolution of human intelligence, McGraw–Hill, New York, p. 239, 2002. Return to text.
- Wong, ref. 4, p. 45. Return to text.
- Wong, ref. 4, pp. 45–46. Return to text.
- Henneberg and Thorne, ref. 26, p. 3. Return to text.
- Morwood et al., ref. 3, pp. 1087–1091. Return to text.
- Morwood et al., ref. 3, pp. 1088–1089. Return to text.
- Morwood et al., ref. 48, pp. 6–8. Return to text.
- Morwood et al., ref. 48, p. 12. Return to text.
- Morwood, M.J., O’Sullivan, P.B., Aziz, F. and Raza, A., Fission–track ages of stone tools and fossils on the east Indonesian island of Flores, Nature 392:176, 1998. Return to text.
- Brumm, A. et al., Early stone technology on Flores and its implications for Homo floresiensis, Nature 441:624–628, 2006. Return to text.
- Brumm et al., ref. 60, p. 628. Return to text.
- The term hominid is an evolutionary term meaning humans and their fossil ‘ancestors.’ However, because of a new classification system many evolutionists now use the term hominin instead. The similar term hominoid means humans and their ‘ancestors’ plus great apes. Return to text.
- Brumm et al., ref. 60, p. 624. Return to text.
- Gribbin J. and Cherfas, J., The First Chimpanzee: In Search of Human Origins, Penguin Books, London, p. 59, 2001. Return to text.
- Reader, J., Missing Links: The Hunt for Earliest Man, Second Edition, Penguin Books, London, p. xv, 1988. Return to text.
- Groves, C. Homo floresiensis: reactions and replies, Before Farming (online journal) 4, article 1, p. 2, 2004. Return to text.
- Dalton, R., Little lady of Flores forces rethink of human evolution, Nature 431:1029, 2004. Return to text.
- Hopkin, M., Old tools shed light on hobbit origins, 31 May 2006, nature.com, accessed 1 June 2006. Return to text.
- Kohn, M., The little troublemaker, New Scientist 186(2504):45, 2005. Return to text.
- Rana, F. and Ross, H., Who as Adam?, NavPress, Colorado Springs, CO, p. 177, 2005. Return to text.
- Wieland, C., Soggy dwarf bones, 28 October 2004; creation.com/dwarfbones. Return to text.
- Wieland, C., Hobbling the Hobbit, 8 November 2004; creation.com/hobblinghobbit. Return to text.
- Wise, K., The Flores skeleton and human baraminology, Occasional Papers of the BSG No. 6, p. 3, 2 November 2005. Return to text.



















Readers’ comments
Comments are automatically closed 14 days after publication.