
New Dmanisi skull threatens to bring the house down

Details of a remarkable fifth Dmanisi cranium (D4500), recovered from a site in Dmanisi, Georgia containing the ruins of a medieval fortress, were published in the journal Science on 18 October 2013.1 The cranium was recovered in 2005 and, together with a mandible (D2600) found five years earlier, constitutes a complete adult skull (together referred to as simply Skull 5). All five Dmanisi skulls are dated by evolutionists to around 1.8 million years ago (Ma), and it is said that “the five skulls were found close together physically, and they were all deposited within a few hundred years of each other in what had been a cave, now collapsed.”2
According to geo-archaeologist Reid Ferring, who dated the site, all “five individuals were found in underground dens where carnivores had probably dragged their carcasses”, with the dens later collapsing.3 The cranial capacity of 546 cubic centimeters (cc) for Dmanisi Skull 5 is the smallest of the Dmanisi sample, with cranial capacities of skulls 1 to 4 reported to be between 601 to 730 cc.4
In 2003 it was reported that the D2600 “mandible is far too large to fit comfortably with any of the crania yet discovered.”5 Hence, expecting to unearth a huge cranium to go with the massive D2600 mandible (lower jaw), one of the surprises was the fit of the enormous D2600 mandible with the smallest-brained Dmanisi crania (D4500).3 The researchers, led by David Lordkipanidze, stated that:
“D4500/D2600 combines a small braincase (546 cubic centimeters) with a large prognathic face and exhibits close morphological affinities with the earliest known Homo fossils from Africa. The Dmanisi sample, which now comprises five crania, provides direct evidence for wide morphological variation within and among early Homo paleodemes. This implies the existence of a single evolving lineage of early Homo, with phylogeographic continuity across continents.”6
The analysis of the Dmanisi skulls showed that:
“The skulls were as variable as African fossils traditionally classified in three different species—H. erectus, H. habilis, and H. rudolfensis. If the Dmanisi fossils had been found in separate places in Africa, they could have been called separate species, Ponce de León says. Lumping them all into H. erectus suggests that the early Homo fossils in Africa may also belong to that same, single lineage.”7
What appears to be essentially suggested by the authors of the new study is that Homo erectus (including versions such as Homo ergaster), Homo habilis and Homo rudolfensis were all members of the same species, and that this single species evolved for a while as a single lineage, later giving rise to subsequent members of the genus Homo. Whilst the new skull (Skull 5) got much of the usual publicity and accolades associated with any alleged ‘hominid’ fossil find that can be construed as providing support for the idea of human evolution, the ‘single evolving lineage’ interpretation of the findings by Lordkipanidze et al. did not endear them to everyone, particularly to splitters. The latter being paleoanthropologists who believe that several distinct co-existing species of hominids gave rise to multiple evolving lineages. In fact, this single lineage idea has been described as “setting off a small ‘bomb’ in the field,” according to co-author Philip Rightmire.7 Perhaps worried about the disarray the single lineage idea will bring to the field of human evolution, one of the doyens of paleoanthropology, Bernard Wood, is quoted by Brian Switek as arguing “that it’s unreasonable to ‘bring the whole bloody house down’ by lumping all early human fossils into a single lineage.”8 A single lineage model would certainly require a major overhaul of all textbooks, museum displays, etc.
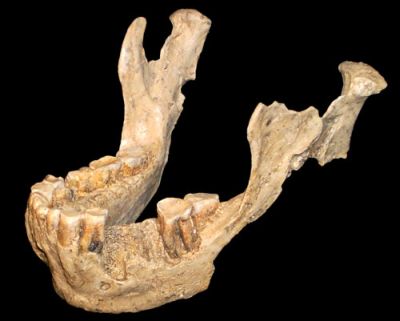
Rather than lumping Homo habilis and Homo rudolfensis with Homo erectus, I have previously argued that they should instead be lumped in with the australopithecines, possibly as the same species as Australopithecus sediba, and I see no reason to change this assessment because of Skull 5.9 One reason is that one also has to consider the rest of the skeleton (the postcranial remains). Whilst there are no postcranial remains reliably linked to Homo rudolfensis,10 the only set of postcranial remains linked with Homo habilis sensu stricto that are associated with “taxonomically diagnostic cranial elements” is OH 62, and its estimated limb-length proportions are as ape-like as Australopithecus afarensis.11 In fact, one evolutionist has argued that although “living nearly a million and a half years after Lucy, the OH62 animal was more ape-like in form than she.”12 As for the Dmanisi specimens, an earlier study by Lordkipanidze et al. described the Dmanisi postcranial remains as having “derived features” that “include modern-human-like body proportions and lower limb morphology indicative of the capability for long-distance travel.”13 That the “Dmanisi individuals appear to have long legs and short arms based on the fossils that have been found,”14 is very different from the “relatively long arms still adapted for climbing in Homo habilis.”8
Also, despite Skull 5 being described as having “had a number of primitive features: a long apelike face, large teeth and a tiny braincase,”15 the “skull’s vertically orientated upper face and the shape of the braincase distinguish it from Australopithecus.”3 The skull’s braincase has been described as “shaped like a typical H. erectus despite its small size.”16 Paleoanthropologist Fred Spoor argues the methods of analysis used by the team in the new study were not sufficient to infer that fossils from Homo erectus, Homo habilis and Homo rudolfensis were of the same species, as they did “a very general shape analysis of the cranium which describes the shape of the face and braincase in broad sweeping terms,” and according to Spoor the “problem is that those Homo species are not defined using such a broad overview of what their general cranial shape is.”17
According to Melissa Hogenboom, Spoor adds “that the very specific characteristics that had been used to define H.erectus, H.habilis and H.rudolfensis ‘were not captured by the landmarks that they use’.”17 Spoor is further quoted as saying that the team “did not consider that the thick and protruding brow ridges, the angular back of the braincase and some details of the base of the cranium are derived features for H.erectus, and not present in H.habilis and H.rudolfensis.”17 Also, in another news article, Spoor “points out that Lodkipanidze’s analysis suggests even the much more ape-like hominins in the genus Australopithecus belong to the H. erectus group. It is not surprising then, that the new analysis misses the more subtle shape differences between Homo species.”16 Hence, it may well be that the similarities between the new Dmanisi Homo erectus skull and Homo habilis and Homo rudolfensis are not as close as is perhaps suggested.
So, how to explain the Dmanisi fossils? As discussed in an upcoming paper dealing with robust humans (Neandertals, Homo heidelbergensis and Homo erectus),18 written before the announcement of this latest find, if the LB1 Homo floresiensis cranium, most recently estimated to be 426 cc,19 belonged to a pathological robust human with cretinism, it raises interesting questions about similar pathology in other small-brained robust humans, such as the Dmanisi Homo erectus specimens. In this context it should be noted that a recent study using geometric morphometric comparative analysis reported that the sole LB1 Homo floresiensis cranium “shows particular affinities in neurocranial shape with the Dmanisi hominins”.20 It would not be that surprising if some of the alleged hominids were instead robust humans that had suffered from cretinism, given that many features of cretinism mimic so-called ‘primitive’ features of evolution. According to evolutionary paleoanthropologist and anatomist Charles Oxnard:

“It is remarkable that so many features similar to those normally present in great apes, in Australopithecus and Paranthropus, and in early Homo (e.g., H. erectus and even to some degree, H. neanderthalensis) but not in modern H. sapiens are generated in humans by growth deficits due to the absence of thyroid hormone. In other words, many of the pathological features of cretinism mimic the primitive characters of evolution making it easy to mistake pathological features for primitive characters. The differences can be disentangled by understanding the underlying biology of characters.”21
If a ‘modern’ human with cretinism can have many pathological features that mimic the so-called ‘primitive’ features of evolution, it is highly likely that a ‘robust’ human with cretinism will have as many, if not even more such features. Cretinism may also explain the large morphological variability in the Dmanisi sample. That is because the bones of cretins are enormously variable, “as would be expected in a pathology with different degrees of affect, and conflation with associated conditions”.22 It is interesting that a lack of chin and a “large jaw compared with the rest of the skull” has been documented in one cretin skull (Dolega),23 as a large jaw (compared with the cranium) is a feature of Dmanisi Skull 5. That the Dmanisi specimens are found in the same locality may not be that unusual. For example, and from an evolutionary perspective, Oxnard suggests that in “seasonally mobile hunter-gatherer groups,” in prior times, cretin children would:
“… be ostracised as adults by the wider community due to their abnormal features and behaviours. Unable to travel easily with a mobile community, especially unable to help build normal temporary dwellings in such a community, adult cretins might well separate and shelter in caves. If there were a reasonable number of them (say, conservatively) 5% of all births, they might indeed shelter together.”24

Alternatives to the above scenario are certainly possible, particularly as there is evidence that early people at least on occasion cared for the infirm. Maybe the cretins were cared for as a group by healthier members of the small, isolated group. As the Dmanisi specimens were located close together physically, and appear to have lived at the same time, it is possible that, as cretins sheltered as a group in a cave, they may have met some unsavory fate together, such as being caught in a cave collapse, or some other event. The above scenario is very speculative, and it may well be that the considerable morphological variation, as well as the extremely small brain sizes, evident in the Dmanisi Homo erectus population, were within normal limits of variation for robust humans, although I consider this option less likely.
If iodine deficiency disorders like cretinism are still a problem in some parts of the world today, despite modern medicine and information about iodine deficiency at our disposal, how much more of a problem could it potentially have been for early post-Flood/post-Babel human populations migrating to uncharted regions of the earth, most likely unaware of the problem (or cause of the problem)—and probably having their hands full just surviving day-to-day? Hence, robust human populations settling in any iodine-deficient regions of Africa, Georgia, China, Indonesia, etc. may well have had a high incidence of cretinism. The Dmanisi population were most likely derived from early post-Babel migrations, and given the difficult situations faced by such early settlers, it is not surprising that only simple stone tools are associated with the finds, said by evolutionists to be “manufactured according to much the same primitive Oldowan tradition that hominids in Africa were practicing nearly a million years earlier.”25 However, it may instead indicate that when these early settlers moved on they took their more sophisticated tools and items with them, and/or that they did not stay in the local area for any length of time, or some other explanation. We just do not know enough about these people to fill in all the blanks.
References
- Lordkipanidze, D., Ponce de León, M.S., et al., A complete skull from Dmanisi, Georgia, and the evolutionary biology of early Homo, Science, 342:326–331, 2013. Return to text.
- Lemonick, M.D., Rethinking your relatives—the fossilized ones, Time, 17 October 2013, www.time.com. Return to text.
- Gibbons, A., Stunning skull gives a fresh portrait of early humans, Science, 342:297–298, 2013; p. 297. Return to text.
- Lordkipanidze et al., ref. 1, p. 327. Return to text.
- Wong, K., Stranger in a new land, Scientific American, 289(5):74–83, November 2003; p. 76. At the time crania D2700, D2280 and D2282 were described in the literature and Wong’s article. The fourth Dmanisi caranium (D3444) is said to have been recovered during excavations in 2002/2003, and was described in 2006 (see Lordkipanidze, D. et al., A fourth hominin skull from Dmanisi, Georgia, The Anatomical Record Part A, 288A: 1146–1157, 2006; p. 1147). Return to text.
- Lordkipanidze et al., ref. 1, p. 326. Return to text.
- Gibbons, ref. 3, p. 298. Return to text.
- Switek, B., Beautiful skull spurs debate on human history, National Geographic, 17 October 2013, www.nationalgeographic.com. Return to text.
- Line, P., Sorting early Homo, Journal of Creation, 27(1):13–15, 2013. Return to text.
- Wood, B., Reconstructing human evolution: achievements, challenges, and opportunities, Proceedings of the National Academy of Sciences, 107(suppl. 2): 8902–8909, 2010; p. 8905. Return to text.
- Wood, B. (Editor), Wiley-Blackwell Encyclopedia of Human Evolution, Wiley-Blackwell, West Sussex, UK, pp. 568–569, 2011. Return to text.
- Stanley, S.M., Children of the Ice Age: How a Global Catastrophe Allowed Humans to Evolve, W.H. Freeman and Company, New York, p. 196, 1998. Return to text.
- Lordkipanidze, D., Jashashvili, T., et al., Postcranial evidence from early Homo from Dmanisi, Georgia, Nature, 449:305–310, 2007; p. 310. Return to text.
- Landau, E., Rare skull sparks human evolution controversy, CNN, 17 October 2013, www.cnn.com. Return to text.
- Wilford, J.N., Skull fossil suggests simpler human lineage, New York Times, 17 October 2013, www.nytimes.com. Return to text.
- Marshall, M., Complete skull of 1.8-million-year-old hominin found, New Scientist, 17 October 2013, www.newscientist.com. Return to text.
- Hogenboom, M., Blow to multiple species idea, BBC, 17 October 2013, www.bbc.co.uk. Return to text.
- Line, P., Explaining robust humans, Journal of Creation, (in press), 2013. Return to text.
- Kubo, D., Kono, R.T. and Kaifu, Y., Brain size of Homo floresiensis and its evolutionary implications, Proceedings of the Royal Society B, 280: 20130338, 2013. http://dx.doi.org/10.1098/rspb.2013.0338. Return to text.
- Baab, K.L., McNulty, K.P. and Harvati, K., Homo floresiensis contextualized: a geometric morphometric comparative analysis of fossil and pathological human samples, PLoS ONE, 8(7): e69119 (p. 9), 2013, doi:10.1371/journal.pone.0069119. Being published before the announcement of Skull 5, this study did not include Skull 5 in the analysis. Return to text.
- Oxnard, C., Ghostly Muscles, Wrinkled Brains, Heresies and Hobbits, World Scientific, Singapore, p. 342, 2008. Return to text.
- Oxnard, C., Obendorf, P.J., Kefford, B.J. and Dennison, J., More on the Liang Bua finds and modern human cretins, HOMO—Journal of Comparative Human Biology, 63:410, 2012. Return to text.
- Oxnard, ref. 21, pp. 320–321. Return to text.
- Oxnard, ref. 21, pp. 339. Return to text.
- Wong, ref. 5, p. 78. Return to text.



Readers’ comments
Comments are automatically closed 14 days after publication.