

Making sense of ‘Homo naledi’
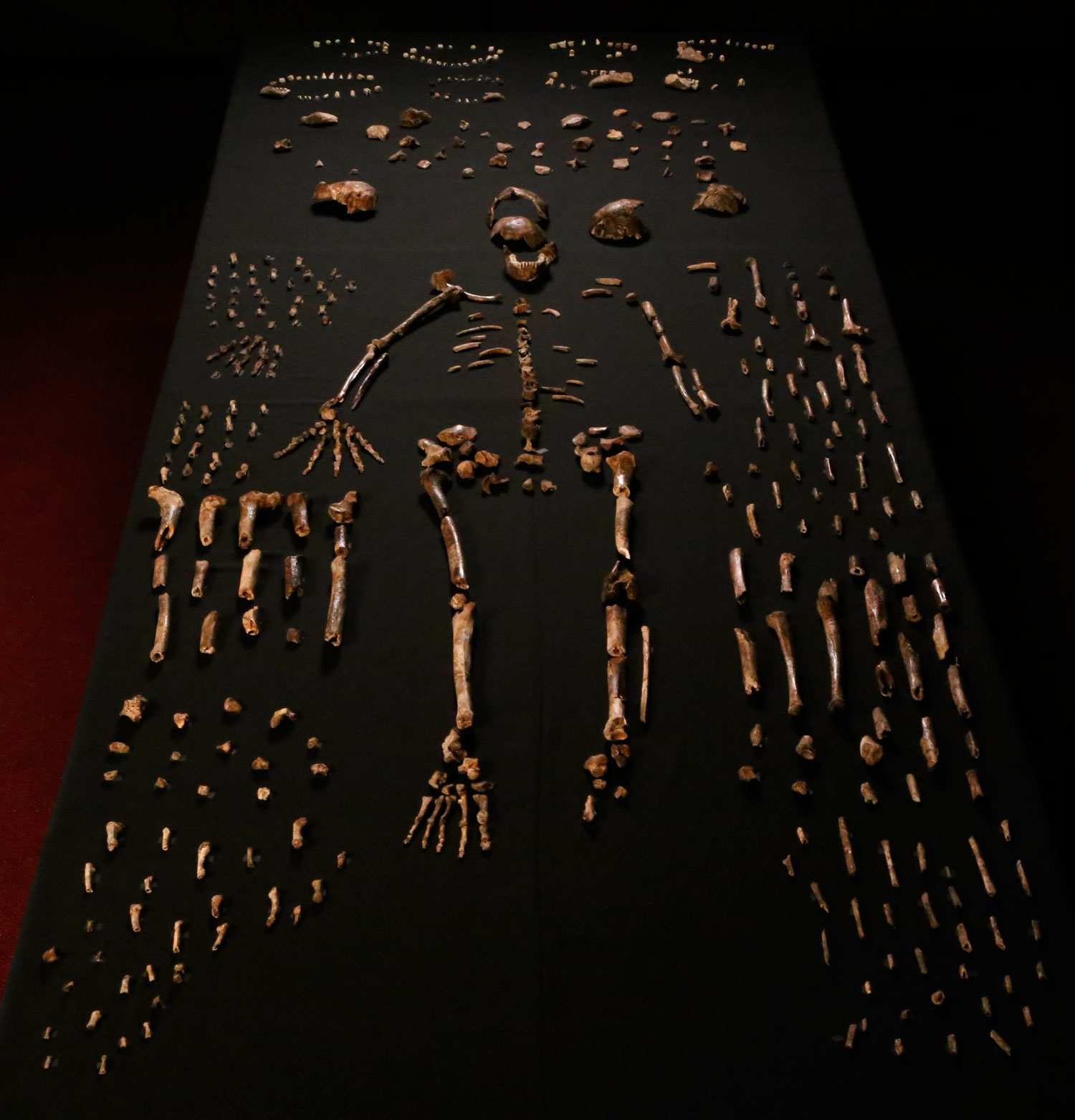
In the field of human origins, a 2015 announcement caused a sensation. It concerned the discovery in South Africa of fossil remains named Homo naledi, by Lee Berger and his team.1 Found in the Rising Star cave system, in areas which had extremely difficult access, the remains were seemingly those of individuals who had climbed in and become trapped—or who were already dead, deposited there by living humans. It is hard to imagine apes doing the trek, particularly in total darkness, nor of humans making the arduous journey just to discard animal remains. Scenarios positing water deposition seem implausible, and there is no evidence of a mixture of human and ape bones. Hence, the context of the find suggests human remains. (A more detailed and extensively referenced article of mine on this topic is available online.2)
Puzzling for all sides
Homo naledi had an essentially modern human ankle and foot (e.g. non-opposable big toe), as well as long lower limbs, indicating it walked upright like humans. The hand and wrist are also like modern humans, except the fingers display a strange curvature; particularly the intermediate phalanges. These middle finger bones are more curved than those of chimpanzees and australopithecines; the median value of their curvature is even higher than for orangutans. As its feet and legs are not built for tree-dwelling, this was unexpected.
Homo naledi is said to display features like our alleged ancestors, the australopithecines (e.g. Lucy), including a pyramidal-shaped (triangular) thorax/ribcage (unverified), a small brain, and a lateral (sideways) flaring of the ilium (the broad upper part of the hip bone in each half of the pelvis). However, a similar lateral iliac flare is also found in specimens of Homo heidelbergensis (SH Pelvis 1) and the Neandertals (Kebara 2),3 that are both undoubtedly human groups. The same iliac flare is found in the hip bone (LB1/7) of Homo floresiensis (dubbed ‘the Hobbit’), which some evolutionists regard as a pathological human.3 However, in all of these, the iliac blade surface is said to be curved, not flat as in some australopithecines (e.g., Australopithecus afarensis, as represented by Lucy) and Homo naledi.3 Lateral flare has also been documented in a pelvis from a modern human, which was said to approach “australopithecine morphology”.4 Just as confusing, it has been reported that a pelvis of a skeleton (KSD-VP-1/1), attributed to afarensis (even though there were no skull and tooth remains to justify this classification), “may have had a more modern human-like orientation of the iliac ala [the large expanded portion of the ilium]”.3 Hence, interpreting a fossil solely on ilium orientation is risky.
As an aside, KSD-VP-1/1 is likely Homo erectus (i.e., human), but as evolutionists say it is about 3.6 million years old, it is assigned to A. afarensis, as admitting H. erectus was around ‘then’ would essentially collapse the human evolutionary story.5
Small brain not diagnostic of non-humans

The smallest estimate of the cranial capacity (correlated with brain size) of the Homo naledi skulls (the DH3/DH4 composite cranium, i.e. made up of two separate specimens) is 465 cc. But this is still larger than the 426 cc of the Indonesian Homo floresiensis LB1, regarded by some evolutionists as a pathological human. And the only cranial capacity estimate of an intact Homo naledi cranium (LES1) is 610 cc, within the lower range of Homo erectus—a group even some evolutionists are saying should be renamed Homo sapiens.
Brain endocast studies use casts of a skull’s interior to get an idea of the shape of the brain it contained. While one can read too much into these, they suggest the brain anatomy of H. naledi was similar to that of humans, not australopithecines.6
Features of H. naledi seemingly outside ‘normal’ variation, both of humans and australopithecines, include the extreme finger curvature, low humeral torsion (the angle by which the orientation of the elbow axis deviates from that of the head of the upper arm bone), and the degree to which the shoulder joint is cranially oriented, i.e. tipped towards the head. These features do, however, illustrate how Homo naledi does not fit as a transitional form between the australopithecines and a supposedly ‘later’ species of Homo—its ‘australopithecine’ features are further from the human condition than in any alleged australopithecine ancestor. Homo naledi does not fit into any evolutionary story.
Are naledi features from cretinism?
When it comes to Homo floresiensis, some evolutionist experts have postulated that these bones represent cretinism in humans. This causes major developmental problems, such as bony deformities, mental retardation, and more. It is caused by iodine deficiency in the unborn. Cretinism is thus common in areas with low environmental iodine, where there is also a high incidence of adult goitre (thyroid enlargement, thus neck swelling, from insufficient iodine).7
In fact, the ‘Cradle of Humankind’ area, where the naledi fossils were found, is located on or near the fringes of what was, prior to iodine supplementation, a narrow ‘goitre belt’, or region in which this deficiency disease was very common. The belt ran 500 km across South Africa’s Transvaal from Zeerust to Nelspruit.8 This regional history increases the likelihood of cretinism in these fossils.
Features mentioned for naledi which are noted in cretinism include triangular thorax, small brain, low humeral torsion,9 and lateral iliac flare.10 Whether it explains the extreme finger curvature and cranially oriented shoulder joint is unclear. If there is a developmental linkage between the shoulder girdle and the upper thorax, then a narrow upper thorax, a feature of cretinism, could potentially explain the shoulder joint being tipped inward towards the head.
Curvature in the fingers is usually caused by mechanical strain from habitual behaviour, such as the high loads experienced when grasping during climbing or moving through the trees suspended by the hands. However, the curvature could also be caused by habitual tool use, or some other repetitive use of the hands, particularly if the bone was more flexible because of pathology (as for example in the rickets caused by vitamin D deficiency).
Interestingly, the proximal phalanges of Homo floresiensis, believed by some evolutionists to involve humans suffering from cretinism, are said to be “curved to a similar degree as in Au. [Australopithecus] afarensis”, and there is evidence that stone tool-making and use were part of this population’s “behavioural repertoire”.11 Hence, frequent stone tool making and use in individuals with cretinism might explain the finger curvature seen in both Homo floresiensis and Homo naledi.
Prior to obtaining an ‘age’, Hawks and Berger considered Homo naledi more “primitive” than Homo erectus, which meant “the H. naledi lineage must have existed earlier than the first occurrence of H. erectus around 1.8 Ma [million years ago].”12 When H. naledi was subsequently ‘dated’ to be much younger than anticipated, between 236 and 335 ka (thousand years ago),13 it seemingly sidelined naledi to being a mere curiosity on the evolutionary storyline.
As an interesting aside from p. 26 of the same (Dirks et al.) paper, two bone fragments were radiocarbon dated to c. 33.0 ka and 35.5 ka. This illustrates the ambiguity of dating methods. It also suggests that the Homo naledi remains may be much more recent than proposed.
Amazingly, one prominent paleoanthropologist “thinks H. naledi branched off from other humans relatively recently and then evolved to look more primitive.”14
Such mental gymnastics further highlight that rather than being an example of ape-man transition, naledi’s peculiar mix of features is puzzling to evolutionists. A more plausible explanation would be as follows.
Explaining Homo naledi
These were post-Fall descendants of Adam, perhaps erectus-like post-Babel humans—an erectus-like cranium (SK 847) has been found at Swartkrans, about 800 metres from the naledi site.15 A number of them suffered from a developmental pathology, cretinism, as suggested by the region’s history of endemic iodine deficiency. According to distinguished evolutionist Charles Oxnard, “many of the pathological features of cretinism mimic the primitive characters of evolution making it easy to mistake pathological features for primitive characters.”16
Cretinism is thus at least a working hypothesis for these remains. It would explain why they have posed a conundrum for evolutionists. And if the spurious evolutionary ‘dates’ are ignored, it would show how here, too, the facts comfortably fit biblical creation.
References and notes
- Berger, L.R. et al., Homo naledi, a new species of the genus Homo from the Dinaledi Chamber, South Africa, eLife 4:e09560, 2015 | doi:10.7554/eLife.09560. Return to text.
- Line, P., Den of ape-men or chambers of the sickly? An update on Homo naledi, 25 May 2017. Return to text.
- VanSickle, C. et al. Homo naledi pelvic remains from the Dinaledi Chamber, South Africa, Journal of Human Evolution, p. 11, 2017 | doi:10.1016/j.jhevol.2017.10.001. Return to text.
- Lovejoy, C.O. et al., The Gait of Australopithecus¸ American Journal of Physical Anthropology, 38:761, 1973. Return to text.
- Line, P., He ain’t my brother: no apparent family ties between Big Man and Lucy, 23 September 2010. Return to text.
- Holloway, R.L. et al., Endocast morphology of Homo naledi from the Dinaledi Chamber, South Africa, PNAS, 14 May 2018 | doi:10.1073/pnas.1720842115. LES1 was not part of study. See also: Wits University, Where hominid brains are concerned, size doesn’t matter, phys.org, 14 May 2018. Return to text.
- Pharoah, P., et al. Endemic Cretinism. In: Stanbury, J.B. and Hetzel, B.S. (Eds.), Endemic Goiter and Endemic Cretinism, John Wiley & Sons, Inc., New York, pp. 410–415, 1980. Return to text.
- Kelly, F.C. and Snedden, W.W., Prevalence and geographical distribution of endemic goiter, in: World Health Organization Monograph Series No. 44, Endemic Goitre. WHO, Geneva, pp. 116, 145, 1960. Return to text.
- Obendorf, P.J. et al. Are the small human-like fossils found on Flores human endemic cretins? Proceedings of the Royal Society B, 275:1289–1290, 2008 | doi:10.1098/rspb.2007.1488. Return to text.
- Oxnard, C., Ghostly Muscles, Wrinkled Brains, Heresies and Hobbits, World Scientific, Singapore, p. 319, 2008. Return to text.
- Kivell, T.L., Evidence in hand: recent discoveries and the early evolution of human manual manipulation, Phil. Trans. R. Soc. B 370(682):20150105, p. 6, 2015 | doi:10.1098/rstb.2015.0105. Return to text.
- Hawks, J. and Berger, L.R., The impact of a date for understanding the importance of Homo naledi, Transactions of the Royal Society of South Africa, 71(2):127, 2016. Return to text.
- Dirks, P.H.G.M. et al., The age of Homo naledi and associated sediments in the Rising Star Cave, South Africa, eLife 6:e24231, p. 34, 2017 | doi:10.7554/eLife.24231. Return to text.
- Barras, C., Meet Neo, the most complete Homo naledi, New Scientist, 234(3125):9, 13 May 2017. Return to text.
- Clarke, R. J. Advances in Understanding the Craniofacial Anatomy of South African Early Hominids; in: Corruccini, R.S. and Ciochon R.L. (Eds.), Integrative Paths to the Past: Paleoanthropological Advances in Honor of F. Clark Howell, Prentice-Hall, Inc., Englewood Cliffs, New Jersey, pp. 216–218, 1994. Return to text.
- Oxnard, ref. 10, p. 342. Return to text.



Readers’ comments
Comments are automatically closed 14 days after publication.