The secularized mainstream media (MSM) are gleefully promoting a recent find, Tiktaalik roseae (right), as the end of any creationist or intelligent design idea. Some paleontologists are claiming that this is ‘a link between fishes and land vertebrates that might in time become as much of an evolutionary icon as the proto-bird Archaeopteryx.’1
So is Tiktaalik real evidence that fish evolved into tetrapods (four-limbed vertebrates, i.e. amphibians, reptiles, mammals and birds)? As will be shown, there are parallels with Archaeopteryx, the famous alleged reptile-bird intermediate, but not in the way the above quote claims!
The above quote comes from two leading European experts in the alleged evolutionary transition from fish–tetrapod, Per Ahlberg and Jennifer Clack. It was about the find of well-known American leaders on the same alleged transition, Neil Shubin and Edward Daeschler, and which was the cover story for Nature.2,3 Clack, Shubin and Daeschler even previously featured on the PBS-Nova seven-part series, Evolution, Episode 2: Great Transformations about the origin of tetrapods.
Shubin et al. found a 20-cm-long skull sticking out of a cliff. They found that this skull, superficially like a crocodile’s, was part of a fish that had a fin that was supposedly on the way to becoming a tetrapod limb. They ‘dated’ it to 383 Ma (million years ago). Since it was in Ellesmere Island, Nunavut Territory (Canada), it was given a genus name from the indigenous Inuktitut word for burbot, or large, shallow freshwater fish.
Is it transitional?
Clack and others are naturally enthusiastic about Tiktaalik’s transitional status. But this is not surprising—to her, we are all fishes anyway! She states:
‘Although humans do not usually think of themselves as fishes, they nonetheless share several fundamental characters that unite them inextricably with their relatives among the fishes … Tetrapods did not evolve from sarcopterygians [lobe-finned fishes]; they are sarcopterygians, just as one would not say that humans evolved from mammals; they are mammals.’4
This is reminiscent of University of Kansas paleontologist Larry Martin criticising overly enthusiastic ‘feathered dinosaur’ claims:
‘You have to put this into perspective. To the people who wrote the paper, the chicken would be a feathered dinosaur.’5
Clack also admitted:
‘There remains a large morphological gap between them and digits as seen in, for example, Acanthostega: if the digits evolved from these distal bones, the process must have involved considerable developmental repatterning. …
‘Of course, there are still major gaps in the fossil record. In particular we have almost no information about the step between Tiktaalik and the earliest tetrapods, when the anatomy underwent the most drastic changes, or about what happened in the following Early Carboniferous period, after the end of the Devonian, when tetrapods became fully terrestrial.’1
Indeed, the evolution of land limbs and life on land in general requires many changes, and the fossil record has no evidence of such changes. Geologist Paul Garner writes:
‘[T]here are functional challenges to Darwinian interpretations. For instance, in fish the head, shoulder girdle, and circulatory systems constitute a single mechanical unit. The shoulder girdle is firmly connected to the vertebral column and is an anchor for the muscles involved in lateral undulation of the body, mouth opening, heart contractions, and timing of the blood circulation through the gills.6 However, in amphibians the head is not connected to the shoulder girdle, in order to allow effective terrestrial feeding and locomotion. Evolutionists must suppose that the head became incrementally detached from the shoulder girdle, in a step-wise fashion, with functional intermediates at every stage. However, a satisfactory account of how this might have happened has never been given.’
Indeed, Tiktaalik’s fin was not connected to the main skeleton, so could not have supported its weight on land. The discoverers claim that this could have helped to prop up the body as the fish moved along a water bottom,3 but evolutionists had similar high hopes for the coelacanth fin. However, when a living coelacanth (Latimeria chalumnae) was discovered in 1938, the fins turned out not to be used for walking but for deft manœuvering when swimming.
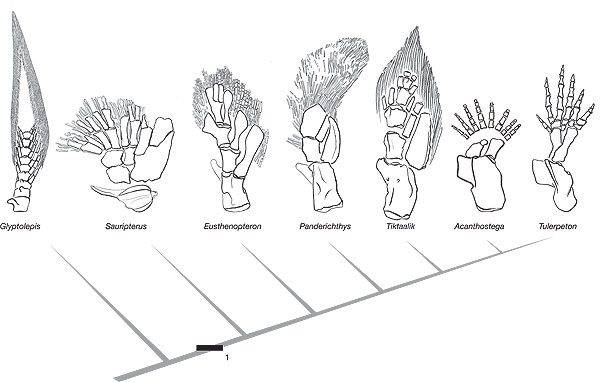
Fig. 2: Cladogram of the pectoral fins on the tetrapod stem, from Ref. 3. Click to see larger image
Quite aside from the huge problems explaining the origin of locomotion, there are other problems. The series of corresponding limbs (Fig. 2, right) does not appear to show the clear progression. Even from looking at it, it is not obvious that the Panderichthys limb belongs in between the adjacent ones in the series. It has fewer small bones. The authors themselves appear to recognize this:
‘In some features, Tiktaalik is similar to rhizodontids such as Sauripterus. These similarities, which are probably homoplastic, include the shape and number of radial articulations on the ulnare, the presence of extensive and branched endochondral radials, and the retention of unjointed lepidotrichia.’
‘Homoplastic’ essentially means ‘convergent’ or ‘analogous’, i.e. independently evolved because of a common function (such as the wings of pterosaurs, bats, birds and insects, according to evolutionists), rather than evolved from a common ancestor (homologous, as evolutionists claim for features such as the different forelimbs here). But homology is alleged to be the evidence for evolution (despite many problems—see Common structures = common ancestry?) But appeal to homoplasy is really explaining away evidence that doesn’t fit the paradigm, and indeed such explaining away is ubiquitous. Two evolutionists admit:
‘Disagreements about the probable homologous or homoplastic nature of shared derived similarities between taxa lie at the core of most conflicting phylogenetic hypotheses.’7
In fact, when more characteristics than just one are analysed, homoplasies become even more necessary to explain away anomalies, as will be explained in the section Mosaic rather than transitional.
Another major problem is that evolutionists appeal to the common pentadactyl 5-digit pattern as evidence for their common ancestry from a 5-digited creature. Yet the nearest creatures they have to a common ancestor did not have five digits! Acanthostega had eight, while Ichthyostega had seven.
Fossil order
Fig. 3: Alleged lineage including Tiktaalik, from ref. 1. Click to see larger image.
Fig. 3 (right) does much to popularize evolution, but there are a number of problems.
The caption admits, ‘These drawings are not to scale, but all animals are between 75 cm and 1.5 m in length.’ If size were taken into account, would there be such a clear progression? Compare a far more extreme example, the supposed land-mammal–to–whale sequence. This was also illustrated as equally sized, but Basilosaurus was 10 times longer than Ambulocetus.
Another admission is, ‘The vertebral column of Panderichthys is poorly known and not shown.’ We should remember the Pakicetus fiasco: when a few bones were known, evolutionists drew it like a half-way land-water form. But when more bones were found, it was realized that it was a fast-running land mammal.
All the fossils of this entire series are assigned to middle-upper Devonian, or 385–365 Ma. Naturally, there are many problems with dating , but even under the evolutionists’ own scenario, there are problems. E.g. the entire fish-to-tetrapod transition is supposed to have occurred in 20 Ma, but other salamanders, according to Shubin himself, have remained unchanged for far longer :
‘Despite its Bathonian age, the new cryptobranchid [salamander] shows extraordinary morphological similarity to its living relatives. This similarity underscores the stasis [no change] within salamander anatomical evolution. Indeed, extant cryptobranchid salamanders can be regarded as living fossils whose structures have remained little changed for over 160 million years.’8
From Wikipedia.org
Fig 4: Lobe-finned fish and amphibians, according to evolutionary order. Click to see larger image.
Even more importantly, the order is not right! Compare Fig. 4 (right): Panderichthys is dated earlier than its supposed predecessor, Eusthenopteron. And all are earlier than the undoubted fish, the coelacanth. This is yet another parallel with alleged bird evolution—undoubted beaked birds like Confuciusornis are 10 Ma older than their alleged feathered dinosaur ‘ancestors’. Evolutionists would argue that it is not a problem, for the same reason that sometimes a grandfather can outlive his grandson. This is correct, but one of the major ‘evidences’ of evolution is how the evolutionary order supposedly matches the fossil sequence. So the mismatch of claimed order of appearance with claimed phylogeny undermines the evolutionary explanation.
Also, Acanthostega is allegedly a predecessor to Ichthyostega, but they were actually contemporaries.
Mosaic rather than transitional
Many of the alleged transitional forms do not have structures in transition from one form to another. Rather, the alleged transitional nature is a combination of fully-formed structures that in themselves are not transitional.9
For example, Archaeopteryx has fully formed flight feathers, an avian lung and an avian braincase (which is why the ‘hoax’ claim is indefensible), but had allegedly reptile features like a tail and teeth. Alleged whale evolution also has a number of ‘modules’, as documented in Walking whales, nested hierarchies, and chimeras: do they exist? These creatures with a mixture of characteristics are called mosaics or chimeras.
Also, who was the predecessor of whom in the case of Acanthostega and Ichthyostega? It depends on which characteristic one looks at: e.g. Ichthyostega’s skull seems more fish-like than Acanthostega’s, but its shoulder and hips are more robust and land-animal–like.10
‘The same sort of reasoning and logic as was used in this article would apply to the fish-to-tetrapod series. In this proposed reptile-to-mammal series, features do not progress consistently. Some organisms towards the mammal end of the series are devoid of certain mammal-like features present in organisms closer to the reptile end of the series. The majority of the hundred-odd traits examined did not progress consistently.’
Lamb’s paper demonstrates this, using Ahlberg’s own table, showing that:
‘For example, Acanthostega, ninth organism in his series, boasts two tetrapod features that are absent in the tenth organism! ’
The same is true of the limb pattern as shown above. This is also consistent with a designer who used ‘modules’ of different characteristics.
A better explanation
When analyzed in detail, the evidence is consistent not with evolution, but with a particular form of intelligent design. But not just intelligent design in the broad sense, which allows for any sort of designer(s), even aliens (such as the Raëlian cult), and even can allow for evolution (Michael Behe, author of Darwin’s Black Box, accepts evolution, for example).
Rather, it supports a particular subset of ID: the biotic message theory, as proposed by Walter ReMine in The Biotic Message. That is, the evidence from nature points to a single designer, but with a pattern which thwarts evolutionary explanations. In this case, the common modules point to one common designer, but evolution is powerless to explain this modular pattern, since natural selection can work only on organisms as a whole. That is, it cannot select for particular head design as such, but only for creatures that have a head that confers superior fitness. But a designer who worked with different modules could create different creatures with different modules, that fit no consistent evolutionary pattern.
Ahlberg, P.E. and Clack, J.A., Palaeontology: A firm step from water to land,Nature440(7085):747–749, 6 April 2006 | doi:10.1038/440747a. Return to text.
Daeschler, E.B., Shubin, N.H. and Jenkins, F.A., Jr, A Devonian tetrapod-like fish and the evolution of the tetrapod body plan, Nature440(7085):757–763, 6 April 2006 | doi:10.1038/nature04639; see abstract. Return to text.
Shubin, N.H., Daeschler, E.B. and Jenkins, F.A., Jr, The pectoral fin of Tiktaalik roseae and the origin of the tetrapod limb, Nature440(7085):764-771, 6 April 2006 | doi:10.1038/nature04637; see abstract. Return to text.
Clack, J.A., Gaining Ground: The Origin and Evolution of Tetrapods, pp. 17–18, Indiana University Press, Bloomington, 2002. Return to text.
Gudo, M. and Homberger, D.G., Functional morphology of the coracoid bar of the Spiny Dogfish (Squalas acanthias): implications for the evolutionary history of the shoulder girdle of vertebrates, 43rd Annual Meeting of the Palaeontological Association, Manchester, 19–22 December 1999; , 4 April 2003. Click on ‘Abstracts’ then ‘Manchester 1999’. Return to text.
Luckett, W.P. and Hong, N., Phylogenetic relationships between the orders Artiodactyla and Cetacea, J. Mammalian Evolution 5(2):130, 1998. Return to text.
Gao, K.-Q. and Shubin, N.H., Earliest known crown-group salamanders, Nature422(6930):424–428, 2003. Return to text.
Wise, K.P., The origin of life’s major groups; in: Moreland, J.P. (Ed.), The Creation Hypothesis: Scientific Evidence for an Intelligent Designer, InterVarsity Press, Downers Grove, pp. 211–234, 1994; esp. p. 227. Return to text.
Comments are automatically closed 14 days after publication.
Feedback Guidelines
Be constructive & courteous. Don't attack individuals, denominations, or other organizations.
Stay on-topic. We're not here to debate matters like eschatology, baptism, or Bible translation.
Links to external sites and articles will be removed from your submission.
Privacy & Content Ownership
Comments become the property of Creation Ministries International upon submission and may be edited for brevity and clarity.
CMI may choose not to publish your comment depending on how well it fits the guidelines outlined above.
By submitting your comment you are agreeing to receive email updates from Creation Ministries International. You may unsubscribe at any time.
CMI records your real name, email address, and country as a sign of good faith. Privacy Policy
If your comment is published, your name will be displayed as ""
Cancel
Accept & Continue
Close
You are leaving CREATION.com
We have supplied this link to an article on an external website in good faith. But we cannot assume responsibility for, nor be taken as endorsing in any way, any other content or links on any such site. Even the article we are directing you to could, in principle, change without notice on sites we do not control.






Readers’ comments
Comments are automatically closed 14 days after publication.