

Journal of Creation 27(3):8–10, December 2013
Browse our latest digital issue Subscribe
Inbreeding and the origin of races

Credit: wikimedia.org
A frequent question asked of creationists is, “Where did the different human races come from?” There are various ways to answer this within the biblical (‘young-earth’) paradigm and many articles and books have already been written on the subject.1 However, I recently thought of a new way to illustrate the origin of different people groups using the 12 Tribes of Israel as an example.
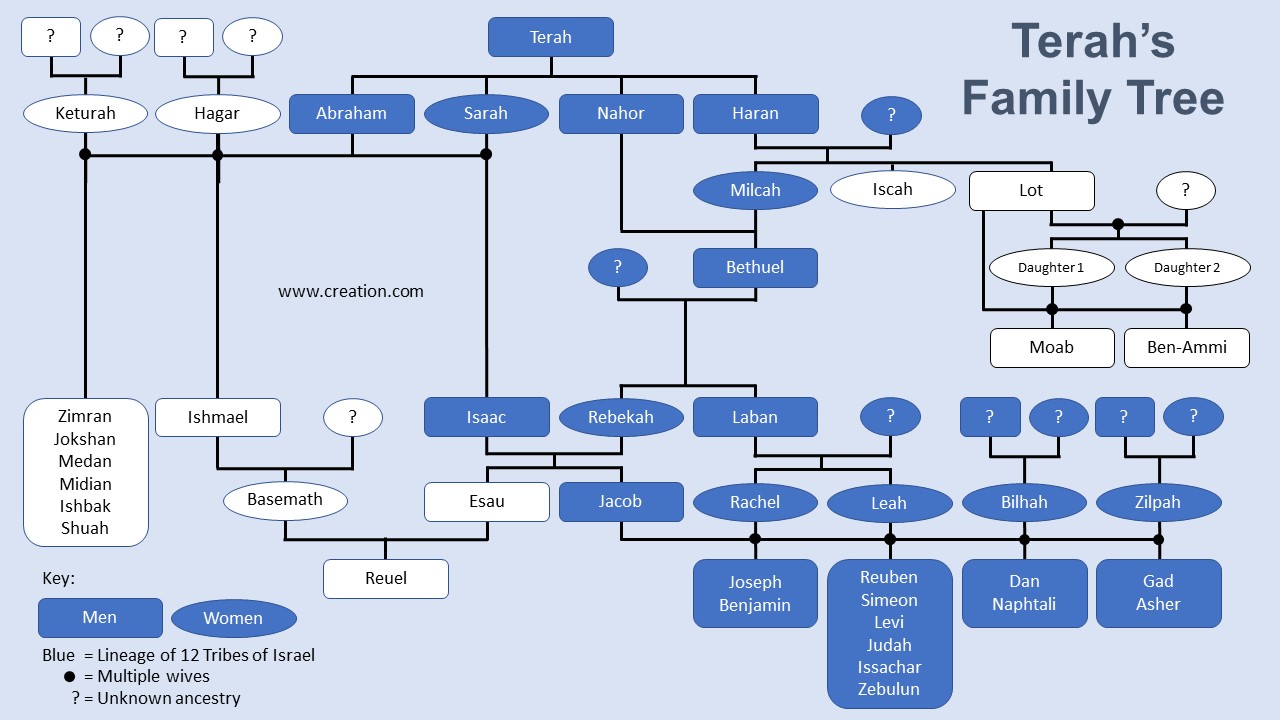
From the Genesis chronogenealogies,2 we have a detailed family history from Noah through Abraham, and then to Jacob and his 12 sons. There is one person in this list, however, who serves as a significant ancestor to the future Jewish nation: Terah, the father of Abram. Terah was also the father of Sarai, though not by Abram’s mother (Genesis 20:12), Nahor, and Haran. It turns out that the four children of Terah are all ancestors of the 12 Tribes of Israel.3 And it is this complex family tree that we can use to illustrate why different people groups across the world don’t all look the same (figure 1).
The science that has rejected ‘race’
When dealing with different people groups, geneticists often calculate a number called the ‘inbreeding coefficient’ and talk about ‘identity by descent’ because every person has inherited multiple identical sections of DNA from both sides of their family. This happens because, if everyone was to trace their family tree back in time, they would eventually come to the same ancestor or set of ancestors in both the mother and father’s side of the family. This can be shown using simple mathematics.
Simply count up the number of ancestors you have in preceding generations: 2 parents, 4 grandparents, 8 great-grandparents, 16 great-great-grandparents, etc., and factor in that the average generation time for modern humans is about 30 years.4 Thus, 10 generations ago was about AD 1710, and you had 1,024 ancestors in that ‘generation’ (Of course, not all of your ancestors lived at exactly the same time, but this is a good estimate.) By 20 generations (about AD 1410), you would have over 1 million ancestors living around that time. At 30 generations (AD 1110), it would seem that you would have over 1 billion ancestors. At least, that is the number of branches that far back in your family tree.
However, since this is probably greater than the world population at that time, it should be clear that there couldn’t be a separate ancestor in each place. And at 40 generations (AD 810) you would have over 1 trillion ancestors, which is impossible since that is more people than have ever lived in the history of the world. Almost all of your ancestors that far back are your ancestors thousands of times over (or more) due to a process I call ‘genealogical recursion’.
Indeed, it does not take many generations to have more ancestral places in your family tree than the population of the world.
The problem is made worse when you consider that many people do not leave any descendants. For example, it is estimated that approximately 25% of the population of Iceland in the early to mid 1800s are the ancestors of about 90% of the modern population, and the results of earlier genealogical work is even more skewed.5 Simply put, the vast majority of ancient people do not have any living descendants. Conversely, Genghis Khan (1162–1227) has about 17 million descendants alive today, including 10% of the 2 million Mongolians and perhaps 0.5% of the world’s population, or 1 out of every 200 people alive today!6
All this means that the calculations in the previous paragraph should be assumed to be maximum times to get to the point where you have more ancestors than the world population at the time, because the effective population size is much smaller that the real population size.
One additional factor to consider is that DNA is inherited in large blocks of unequal size. This is due to the recombination of homologous chromosomes prior to sexual reproduction. My own calculations (data not shown) indicate that in as little as 5 generations (that is, the mid-1800s) you might have people in your family tree from whom you inherited no DNA! This depends on the amount of recombination per generation and the distribution and spacing of recombination events. The various models I have applied all point to a rapid loss of genetic ancestors, irrespective of genealogy, with an average of less than 10 generations to null ancestry for any ancestor-descendant pair in the genealogy. Because Terah’s family tree is shorter than this, the calculations in this paper are probably accurate. That is, we would expect him to be a genetic ancestor to the rest of the people in the genealogy.
The number of ancestors going back in time, coupled with many people not having children forward in time, coupled with the effects of recombination, coupled with the fact that people historically have interbred mostly within their own people group, means that there has been a significant amount of inbreeding in every population.
This is the reason that people from different people groups don’t look the same. Many genes for traits like height, head shape or even eye color don’t necessarily show strong inheritance patterns. Yet, enough inbreeding has occurred in all people groups that strongly associated traits often exist that can be used to roughly distinguish one set of people from another. This is not always easy to do, however, and one can find almost all traits in almost all populations.7 In fact, when looking at the underlying genes, there are about 10 million variations in the human genome that are found in all world populations.8 These are not found at the same frequency in all populations, however, and this gives rise to some of our ideas about ‘races’. We have to be careful, however, when trying to assign ‘race’ to a person, for modern geneticists have rejected the entire notion. As geneticist Lluis Quintana-Murci recently said:
“But the genes that explain the phenotypic differences between populations [i.e., the differences in the way people from different populations look] only represent a tiny part of our genome, confirming once again that the concept of ‘race’ from a genetic standpoint has been abolished.” 9

Israel as a case study
Now, turning to the Bible and to one particular family, that of Israel, we can see these ideas at work. Most people with some knowledge of the Bible know that Abram married his half-sister Sarai, Isaac married his cousin Rebekah, and Jacob married his cousins Rachel and Leah, who gave rise to 8 of the 12 Tribes of Israel. But all these people also descend from Terah. Does this mean that a large fraction of Terah’s DNA should be represented in the 12 tribes due to inbreeding?
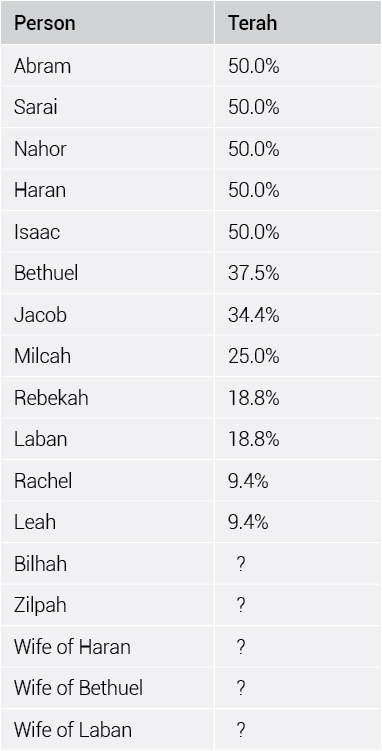
Children each inherit half of the DNA carried by each parent. Thus, a child will carry a quarter of each grandparent’s set of DNA, one-sixteenth of each great-grandparent’s DNA, etc. Abram was 50% identical to Terah. So was his half-sister Sarai. Their child Isaac should have been 25% of each grandparent, but because his parents were both children of Terah, Isaac was 50% identical to his one grandfather (obtaining 25% of Terah’s genome from each parent). Because of inbreeding, he had a similar genetic status as a child of Terah.
The only difference is that Isaac would have inherited identical copies of parts of some chromosomes from his parents. Usually, a person inherits two versions of the human genome, one from the mother and one from the father. In Isaac’s case, however, some of that DNA inherited from Sarai was identical to that inherited from Abram, because they had, in turn, inherited identical parts of Terah’s genome. In those places (about 25% of his genome), any recessive traits were fully expressed.
In Israel’s family tree, multiple generations intermarried. The genealogy is complex. How can one calculate the amount of inbreeding? By simply listing each line from Terah to the 12 sons of Israel and figuring that ½ of Terah’s DNA is lost each generation (table 1).
Using this method, one can generate a percent similarity to Terah for any person in the genealogy (table 2). All one has to do is sum the results for each line that traces back to Terah.
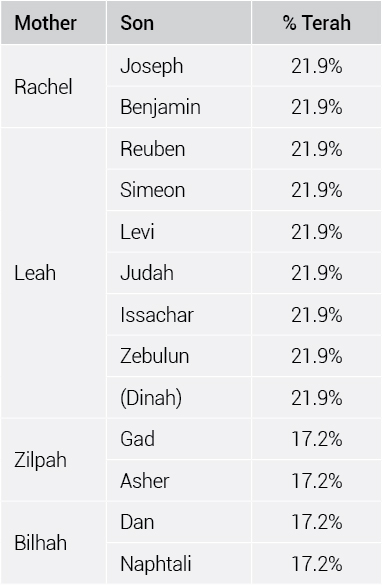
It is also possible to do this for the 12 sons of Israel. For example, Joseph and Benjamin (their mother was Rachel) go back to Terah along lines A, B, C, D, E, and G. But Gad and Ashur (their mother was Bilhah) only go through lines A, B, C, and D. If you do this for each line and sum the results, eight of the 12 sons of Israel (those descended from Leah and Rachel) inherited 21.9% of Terah’s DNA, at a minimum (table 3). After that many generations, they should have been between 6.3% and 1.6% identical to Terah, depending on which line you are examining. The other four (those descended from Zilpah and Bilhah) were 17.2% Terah, instead of 6.3%. Inbreeding makes a huge difference!
Thus, each of the 12 brothers was about 20% genetically identical to their simultaneous fourth, fifth, and sixth-generation ancestor, Terah. This is about as much as you would have expected of grandchildren, not great-great-great-great-grandchildren. These are minimum figures because we do not know the genealogies of many of the wives, but it illustrates quite clearly the degree of inbreeding in this one ancient family.
In fact, we should expect patriarchal societies like this to be inbred, and there are other indications in the Bible. For example, the endemic gigantism in various groups living in Canaan prior to the Israelite conquest10 may have been due to a concentration of some gene(s) associated with gigantism in the population through inbreeding. There is also evidence of polydactyly (extra fingers and toes, a recessive mutation that generally only appears due to inbreeding) in one of the giants killed in David’s time (Numbers 13:28), and there is abundant testimony of left-handedness in the tribe of Benjamin (was this training or genetics?).11
Of course, the Israelites did not remain isolated from the people with whom they lived, and marriages across cultural lines are recorded many times in the Old Testament (e.g. Rahab and Ruth, but marriages with many other non-Israelites are also mentioned). Also, Terah and the other Patriarchs were the ancestors of many other peoples. Thus, there are no ‘Jewish’ genes. They became a ‘Middle Eastern’ people group because they descend from and intermarried with the other people in the area.
The ‘12 Tribes of Israel’ scenario of inbreeding is an excellent example of what we expect to occur thousands of times across the world as people spread out from Babel, following population stratification resulting from the confusion of languages (Genesis 11:1–9). They are not exceptional in the level of inbreeding demonstrable from their family tree, nor are they exceptional in the degree of outbreeding with the closely and distantly related peoples in their area. They are, however, a mirror of the development of the many different people groups on the earth today, and the only reason any ‘racial’ features exist in a population (and it is not difficult to think of many different traits associated with people across the world) is due to inbreeding in those populations in historical times.
References and notes
- See Racism: questions and answers; creation.com/racism-questions-and-answers. Return to text.
- A ‘chronogenealogy’ is different from a ‘genealogy’ in that the former lists specific dates and can be used to build not only a family tree but also a detailed history with dates for specific historical events. The genealogies in the New Testament (cf. Matthew 1 and Luke 3) are simply lists of names. The chronogenealogies in the book of Genesis, however, contain not only names but also the age of each father when the next son was born. It is this that allows us to calculate the age of the earth, etc. See Sarfati, J., Biblical chronogenealogies, J. Creation 17(3):14–18, 2003; creation.com/biblical-chronogenealogies. Return to text.
- The names can get a little confusing. Abraham and Sarah were called Abram and Sarai until God changed their names later. Likewise, God changed Jacob’s name to Israel. I’ve tried to be consistent and I have tried to avoid other words people often substitute for ‘Israelite’, e.g. ‘Jew’, ‘Hebrew’, or even ‘Semite’. We get the phrase ‘Jewish’ from a significant Israelite tribe, Judah, the word ‘Hebrew’ from a significant person in the line of Abraham, Eber, and the word ‘Semitic’ derives from Noah’s son, Shem. Thus, when discussing the 12 sons of Jacob for this paper, ‘Israel’ is really the most precise term to use. See also Sarfati, J., A brief history of the Jews, creation.com/jews, 16 May 2000. Return to text.
- The formula is N = 2n, where N = number of ancestors and n = the number of generations back. The figure “30 years per generation” comes from several studies, including Helgason, A. et al., A populationwide [sic] coalescent analysis of Icelandic matrilineal and patrilineal genealogies: Evidence for a faster evolutionary rate of mtDNA lineages than Y chromosomes, American J. Human Genetics 72:1370–1388, 2003. Even chimpanzees have a longer generation time than the ‘20 years’ normally assumed in prior evolutionary studies, e.g. Langergraber, K.E. et al., Generation times in wild chimpanzees and gorillas suggest earlier divergence times in great apes and human evolution, Proc. Nat. Acad. Sci. USA 109(39):15716–15721, 2012. Return to text.
- Helgason, A. et al., A populationwide [sic] coalescent analysis of Icelandic matrilineal and patrilineal genealogies: evidence for a faster evolutionary rate of mtDNA lineages than Y chromosomes, American J. Human Genetics 72:1370–1388, 2003. Return to text.
- Zerjal, T. et al., The genetic legacy of the Mongols, American J. Human Genetics 72:717–721, 2003. Return to text.
- See Batten, D., A lady of distinction, Creation 25(1):44–45, 2002; creation.com/a-lady-of-distinction. Return to text.
- Carter, R.W., The non-mythical Adam and Eve! Refuting errors by Francis Collins and BioLogos, creation.com/historical-adam-biologos, 20 August 2011. Return to text.
- Quintana-Murci, L., Human variation chalked up to natural selection: study, PhysOrg.com, 4 February 2008. Return to text.
- There are many references, not just to Goliath. For an explanation of why these giant people were not descendants of the ‘Sons of God’ in Genesis 6, see Bates, G., The return of the Nephilim?, creation.com/nephilim, 6 June 2009. Return to text.
- E.g. Judges 3:15–21, 20:16; 1 Chronicles 12:2. Return to text.










Readers’ comments
Comments are automatically closed 14 days after publication.