
What is Homo luzonensis?
Ape or man?
From time to time the media is abuzz with news of yet another human fossil discovery. Each time researchers hope for more clues with which to try to piece together the alleged evolutionary picture of our origins. This time, the fossils come from a part of the world far removed from the usual sites of human fossil research in eastern and southern Africa.
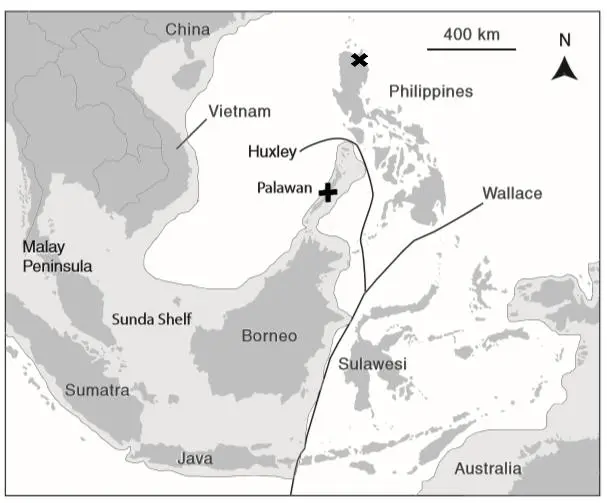
Researchers have found 13 bones, recovered between 2007 and 2015, which are claimed to belong to a new species of hominin, called Homo luzonensis. The species was named after the island of Luzon in the Philippines where it was discovered (see figure 1).
The bones come from three individuals—two adults and a child—in Callao Cave (marked with an X on fig. 1). The bones include a third right metatarsal (bone of the foot), seven post-canine maxillary teeth (from the upper jaw), two manual phalanges (finger bones), two pedal phalanges (toe bones), and a femoral shaft (thigh bone).1 The number of bones is too few to draw any definitive conclusion, but it still may be worth the time to analyze the bones. DNA extraction from H. luzonensis has been unsuccessful.
Evolutionists claim that H. luzonensis lived 67 thousand years ago (ka).2 Obviously, they would like to put H. luzonensis into a timeline of evolutionary development, while creationists would classify such fossil remains as either human or ape-like primate. Where does H. luzonensis fit?
The evidence
Morphology
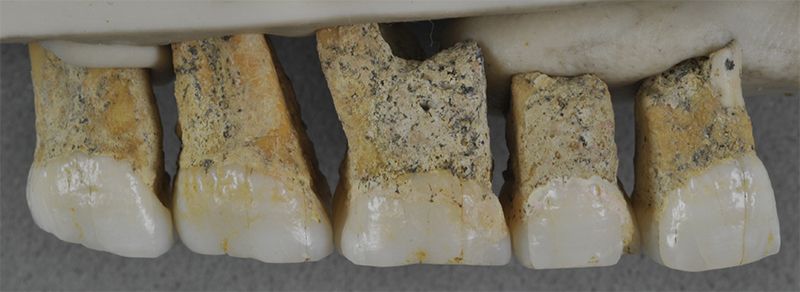
Seven post-canine maxillary teeth (from the upper jaw) were found (five of which are shown in figure 2). The molars are small and follow an M1 > M2 > M3 size pattern3 from front to back, which is characteristic of humans, especially H. floresiensis. The tooth morphology differs from other alleged hominins. The premolars have multiple robust roots, and resemble those of Australopithecus and Paranthropus. However, they also resemble those of Indonesian Homo erectus, which almost all creationists agree is human. Unfortunately, no skull fragments were found.
Two finger bones were found among the fossils. A bone called CCH2 is an intermediate manual phalanx (middle finger bone). This bone is elongated and curved; these are features found in the australopithecines, which are non-human. However, this morphology has also been reported in Homo naledi. A distal manual phalanx (third finger bone), CCH5, has been discovered, which does not articulate with CCH2, the intermediate phalanx, but the proportions of this bone fit into the range of Homo sapiens.
However, H. naledi’s finger curvature can also be explained by modification through use, as in the case of athletes like climbers who do a lot of gripping.4 H. naledi has a relatively long thumb compared to its other fingers, and in other aspects it resembles the hand of modern humans.5,6 H. floresiensis also has curved phalanges,7 yet many creationists classify it, as well as H. naledi, as part of the human baramin. Some suggest that pathology explains some of the strange morphology seen in these fossil ‘species’,8 which may or may not have contributed to the finger curvature.

The third right metatarsal (a bone in the mid-foot) is gracile (slender) and resembles that of H. floresiensis. And it is almost exactly the same length (61.0 mm compared to 60.4 mm for H. floresiensis).2 It fits into the range of other Homo species, and overlaps with the range of variation measured for Philippine Negritos.2 It is different from the same bone in Macaca ( macaques), Hylobates (gibbons) and Pongo (chimps).
Animal remains
90% of the animal remains in the cave are from brown deer (Cervus mariannus), including teeth, and scapular, pelvic and vertebral fragments. A tibia bone from this species with three parallel cut-marks shows evidence of human activity. Remains from the warty pig (Sus philippensis) and an extinct bovid were also found.2 This indicates that H. luzonensis hunted for and processed large-bodied animals for food. This is evidence that luzonensis had a human-like higher intelligence. Also, stone tools associated with a butchered rhinoceros, indicating human intelligence, were found nearby in the Cagayan Valley of northern Luzon, dated by evolutionists to between 777 and 631 ka.9 Whilst not accepting these age dates, they nonetheless indicate there was likely a human presence in the region relatively early post-Flood.
Geography
The island of Luzon lies east of the Huxley Line, which is a modification of the Wallace Line. The Wallace/Huxley lines separate those land areas which were not connected with each other at lower sea levels during the Ice Age.10 Therefore, it is likely that the individuals tagged as ‘H. luzonensis’ would have sailed from mainland Southeast Asia to the island today called Luzon in the modern-day Philippines. Similarly, ‘H. floresiensis’ is believed to have sailed to the island of Flores, which is similarly separated from other islands in Indonesia. Some creationist authors conclude that H. floresiensis would have had to sail across deep-water straits that separated Flores from the continental Sunda Shelf. This is a sign of higher, human intelligence.3
Conclusion
It could be that H. luzonensis is very similar to H. floresiensis and both are human, likely belonging to the Negrito population living in the Philippines. Possibly H. luzonensis, H. floresiensis and the Negritos are all inter-related. It is interesting to note that verified modern human (Homo sapiens) remains have also been found in cave sediments, apparently from well before modern settlement, on the island of Tabon, also in the Philippines (marked with a + on fig. 1). These include a frontal bone as well as two mandibular fragments (parts of the lower jaw).11 Also of interest is the relationship of H. luzonensis to H. erectus and the Denisovans, as well as the small-bodied humans from Palau, Micronesia.12 This is because these humans are also believed to have inhabited South East Asia in the past.
The skeletal remains of H. luzonensis are scant. The bones seem to portray both australopith as well as human characteristics. The teeth of H. luzonensis are human. Though the intermediate phalanx CCH2 may seem ape-like, it is also similar to that of H. naledi, which most creationists think is human. The distal phalanx is similar to that of H. sapiens, meaning that overall, the hand of H. luzonensis is human. The third right metatarsal resembles that of H. floresiensis, also held to be human by most creationists, and is also within the range of Philippine Negritos. All of these bones are either characteristic of humans only or overlap with australopithecines. None of the bones have features which resemble only australopithecines. From the scant evidence there is at this point no reason to think that H. luzonensis is not human.
References and notes
- Détroit, F., Mijares, A.S., Corny, J., Daver, G., Zanolli, C., Dizon, E., Robles, E., et al., A new species of Homo from the Late Pleistocene of the Philippines., Nature. 568(7751):181-186, 2019. Return to text.
- Mijares, A.S., Détroit, F., Piper, P., Grün, R., Bellwood, P., Aubert, M., Champion, G., et al., New evidence for a 67,000-year-old human presence at Callao Cave, Luzon, Philippines, J Hum Evol. 59(1):123-32, 2010. Return to text.
- M1 denotes the first molar, M2 the second, and M3 the third molar from the front of the mouth to the back. Return to text.
- Rupe, C. and Sanford, J., Contested Bones, FMS Publications, 2017. Return to text.
- Kivell, T.L., Deane, A.S., Tocheri, M.W., Orr, C.M., Schmid, P., Hawks, J., Berger, L.R., et al., The hand of Homo naledi, Nat Commun. 6:8431, 2015. Return to text.
- Kivell, T.L., Evidence in hand: recent discoveries and the early evolution of human manual manipulation, Philos Trans R Soc Lond B Biol Sci. 370(1682), 2015. Return to text.
- Larson, S.G., Jungers, W.L., Tocheri, M.W., Orr, C.M., Morwood, M.J., Sutikna, T., Awe, R.D., et al., Descriptions of the upper limb skeleton of Homo floresiensis. J Hum Evol. 57(5):555-70, 2009. Return to text.
- Line, P., Making sense of ‘Homo naledi’, Creation 40(4):36–38, 2018. Return to text.
- Ingicco, T., van den Bergh, G.D., Jago-on, C., Bahain, J.-J, Chacón, M.G., Amano, N., et al., Earliest known hominin activity in the Philippines by 709 thousand years ago. Nature 557:233-237, 2018. Return to text.
- Esselstyn, J.A., Oliveros, C.H., Moyle, R.G., Peterson, A.T., McGuire, J.A., Brown, R.M., Integrating phylogenetic and taxonomic evidence illuminates complex biogeographic patterns along Huxley’s modification of Wallace’s Line, Journal of Biogeography 37:2054–2066, 2010. Return to text.
- Détroit, F., Corny, J., Dizon, E.Z., Mijares, A.S., “Small size” in the Philippine human fossil record: is it meaningful for a better understanding of the evolutionary history of the negritos? Hum Biol. 85(1-3):45-65, 2013. Return to text.
- Berger, L.R., Churchill, S.E., De Klerk, B. and Quinn, R.L., Small-bodied humans from Palau, Micronesia, PLoS ONE 3(3): e1780. doi:10.1371/journal.pone.0001780 Return to text.


Readers’ comments
Comments are automatically closed 14 days after publication.