

Journal of Creation 34(2):55–61, August 2020
Browse our latest digital issue Subscribe
New Homo erectus crania associated with stone tools raise issues

A recent paper describing two new Homo erectus crania from Ethiopia, found directly associated with both Mode 1 and Mode 2 stone tools, raises issues regarding Homo erectus. This article discusses some of these issues, including the stone tools, the small cranial capacity, the robust form of the cranium, and the location and setting in which these fossils were found.
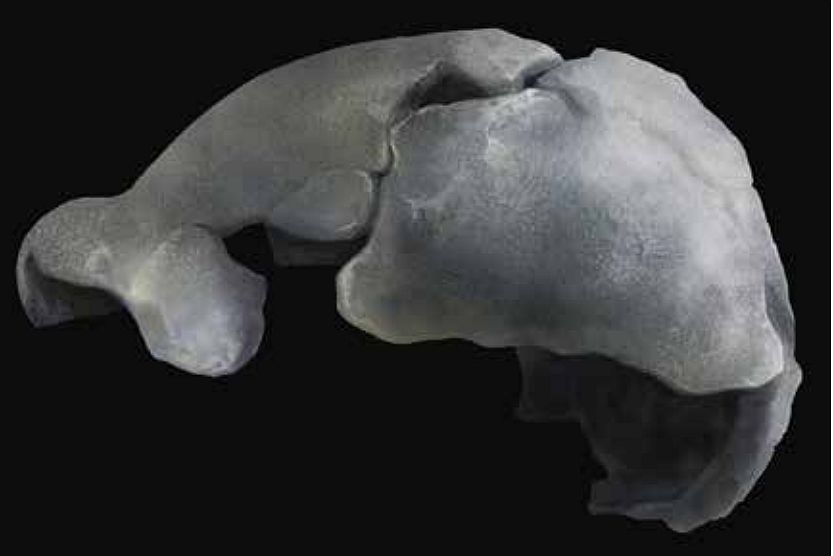
In early March 2020 Semaw et al. published a paper titled “Co-occurrence of Acheulian and Oldowan artifacts with Homo erectus cranial fossils from Gona, Afar, Ethiopia”.1 The paper raises issues regarding Homo erectus that are important to address from a creation point of view. Included in the article is the description of two adult Homo erectus crania from the Gona Project study area, Afar, Ethiopia, neither of which appear to have been published before, even though they were discovered about two decades ago. The larger BSN12/P1 (partial) cranium was discovered in 1999 at the Busidima North site #12, while the smaller DAN5/P1 (relatively well-preserved) cranium (figure 1) was discovered in 2000 at the Dana Aoula North locality #1.2 Although evolutionary (geological) ages of strata and/or fossils are given in this article as information to the reader, I do not accept the validity of these age dates.
Stone tools
An item of interest highlighted by the publication was the direct association of Oldowan and Acheulian stone tools with both crania, these being found about 5.7 km apart, and supposedly separated temporally by at least 240,000 years.3 According to Semaw et al.:
“Here, we report combined Oldowan (Mode 1) and Acheulian (Mode 2) stone tool assemblages and hominin cranial fossils found in direct association that derive from stratigraphic levels dating to 1.26 Ma ago at Busidima North (BSN12) and approximately 1.6 to 1.5 Ma ago at Dana Aoule North (DAN5) … which illuminate H. erectus variability and behavioral flexibility.”4
The authors remarked that it was their observation at Gona and elsewhere that many archaeological sites dating from 0.5 to 1.6 Ma (million years ago) only contained Mode 1 stone tools, and that the Gona evidence suggested “that most of these Mode 1 sites were created by H. erectus (sensu lato), not a different hominin species, particularly in areas that do not preserve evidence of other hominins in the Middle Pleistocene, such as the Afar”.5 They regarded the evidence at Gona as suggesting that Homo erectus “had population-level behavioral diversity and flexibility, with a lengthy and concurrent use of both Mode 1 and Mode 2 technologies”.5
Hence, as indicated above, the finding of a direct association of both simple Oldowan tools and the relatively more complex Acheulian stone tools with Homo erectus crania is strong evidence that humans classified as Homo erectus made and used both types of stone tools (i.e. Modes 1 and 2).
From a creation point of view, where evolutionary age dates are not accepted, this indicates that Homo erectus (not Homo habilis) individuals were also the likely makers of the Oldowan stone tools at Olduvai Gorge, Tanzania, and other places. Homo habilis sensu lato (in a broad sense, e.g. incorporating Homo rudolfensis) have usually been assumed to be the manufacturers of the first known (Oldowan) tools, “even though fossil remains have not usually been found in direct association with the tools”.6 As detailed elsewhere, Homo habilis is likely a phantom species, i.e. a ‘composite taxon’ made up of mainly australopithecines, but also likely a few Homo erectus remains, that have been bundled together and marketed as a species of ‘apeman’.7
There is a caveat to the interpretation of artefacts as Oldowan tools, however. Capuchin monkeys have been observed deliberately breaking stones, unintentionally producing fragments stated as “recurrent, conchoidally fractured, sharp-edged flakes and cores that have the characteristics and morphology of intentionally produced hominin tools”.8 Apparently, as reported by Ewen Callaway, “About half of the flakes made by the capuchins bore the hallmarks of Oldowan tools called choppers, says Proffitt”,9 the latter being the first author of the study. Hence, if Oldowan-like tools could be produced this easily, it makes one wonder whether some of the so-called Oldowan tools were simply accidental by-products of stone breakage, and not necessarily by human hands. Another suggestion, by co-author of the Gona Homo erectus paper, Michael Rogers, as articulated by Ruth Schuster, is that “Possibly the worked stones perceived as Mode I tools were by-products of the manufacturing process of making Mode II implements.”10

Oldowan (Mode 1) tools are basic, essentially “made by smashing two rocks together to form a sharp flake”, whereas Acheulian tools (Mode 2) are more complex, being “made by repeatedly chipping away at a rock to shape it into a hand axe”.11 The Acheulian handaxes (figure 2) are typically teardrop-shaped. They are thought by evolutionists to have first been made by Homo erectus sensu lato, and, according to Stringer and Andrews, “The people who made the handaxes clearly had a specific shape in mind, and often went far beyond a purely utilitarian form in the care with which they produced them.”12 Hence, from a creation point of view, such purposeful manufacture of a tool has the hallmarks of human activity, indicating that Homo erectus individuals were fully human.
Location and setting
Fossilized remains of Homo erectus sensu lato have been discovered in the landmasses of Africa and Eurasia. They appear to be concentrated in several regions, including Java (the central island of Indonesia), China, East Africa (Kenya, Tanzania, Eritrea, Ethiopia), and Georgia (the Dmanisi remains). Remains have also been found in North Africa (Salé 1 cranium from Morocco) and India (Narmada cranium).13 Homo erectus remains have also been found in Denizli, Turkey; namely the partial Kocabaș skullcap.14 Some researchers view the Ceprano 1 calvarium as belonging to Homo erectus, which would also place the species in Italy.15 Ronald Clarke has also suggested that the partial cranium SK 847, from Swartkrans, South Africa, should be classified as an early Homo erectus.16 The two new Homo erectus cranial fossils were from Gona, Afar, Ethiopia, and, according to Semaw et al.:
“Hominin remains and artifacts at DAN5 and BSN12 were discovered in the Busidima Formation, which lies above the well-known Hadar Formation and spans ~2.7 to <0.16 Ma … . The Busidima Formation consists of stacked fining upwards packages of fluvial deposits laid down by a paleo-Awash River.”17
The BSN12/P1 cranium (and associated Mode 1 and Mode 2 artifacts) was found in direct association with the Boolihinan Tuff (BHT) stratigraphic level, where 28 metres “of reversely magnetized sediments overlie the BHT, capped by an additional 18+ m of normally magnetized sediments”.4 The DAN5/P1 cranium and associated artifacts (Mode 1 and Mode 2 tools), found at the DAN5 locality, were said to have
“… come from a siltstone unit in reversed magnetized sediments 6.5 m above the top of the Olduvai Subchron and 10.5 m below the base of the Jaramillo Subchron … , with a well-developed, 8- to 9-m-thick, cumulic paleosol separating the artifacts and cranium from the normally magnetized stratigraphic interval above … .”18
The above setting shows similarities to other Homo erectus fossil finds. In a survey of Homo erectus crania (where cranial capacity had been estimated), the setting in which many of these crania were found was sediments deposited by water.13 With these Homo erectus fossils we are generally not referring to remains that have been deliberately buried, as is the case with some modern human or Neandertal remains; rather crania, and sometimes scraps of other postcranial material, exposed by erosion of the deposits, or otherwise discovered (e.g. by excavation).
For many of these Homo erectus crania (or postcranial remains) to be buried in sediments and subsequently fossilized, and then for the stratigraphic layers of these sediments to later be exposed/eroded, by whatever mechanism, would take time. This is apart from the time it would take for the individuals to migrate to these regions, some of which (e.g. Indonesia) would involve lengthy journeys. Hence, if these fossils were of individuals post-Flood, then it seems likely they represent migrations of people that began very early post-Flood.
In terms of affinity, the more complete and smaller DAN5/P1 cranium is said to show similarity to the Dmanisi crania (1.76–1.85 Ma) from Georgia, the juvenile KNM-ER 42700 cranium (1.5–1.6 Ma) from Kenya, and the small Olorgesailie (KNM-OL 45500) cranium (0.95 Ma) from Kenya, all Homo erectus.19 In regard to the larger and very incomplete BSN12/P1 cranium, it is stated as
“… similar to the more robust African specimens such as Olduvai Hominid 9, the ca. 1-Ma specimens from Middle Awash, Ethiopia [BOU-VP-2/66 …] and Buia, Eritrea [UA-31 …], and those from Indonesia and eastern Asia by having a longer and lower vault with a thickened supraorbital torus.”19
Although the features in Homo erectus crania vary, there are also similarities, even between specimens as far apart as Africa and Indonesia, as indicated above. A possible, logical explanation for this would be that all the Homo erectus individuals from these different regions of the world ultimately trace their origins back to an original and diverse ‘Homo erectus’ population, likely from Babel.
Cranial capacity in Homo erectus
According to Stephen Molnar, the mean cranial capacity for modern humans is 1345 cc (cubic centimetres).20 In comparison, the mean cranial capacity of 48 Homo erectus crania was calculated to be about 934 cc (range of 48 crania was 546 cc to 1260 cc), which is about 70% of the mean cranial capacity in modern humans.13 A smaller body size on average in Homo erectus may account for some of the difference in cranial capacity, but not all. The smaller Gona DAN5/P1 cranium was estimated to have a cranial capacity of ~598 cc, whereas the larger and partial Gona BSN12/P1 cranium was estimated to have a cranial capacity between 800 and 900 cc.19 While the cranial capacity of the BSN12/P1 cranium is close to the mean of Homo erectus, that of the DAN5/P1 cranium is very much at the lower end of the range, having the smallest adult Homo erectus cranial capacity known in Africa. Adding these two crania to the 48 crania above (using the mean value of 850 cc for BSN12/P1 cranium), the mean cranial capacity of 50 Homo erectus crania is about 926 cc. Hence, it may be validly asked why the cranial capacities of all Homo erectus crania are below the modern human mean, some being considerably below. However, this may not necessarily be true.
For example, consider the Xuchang crania, evolutionarily dated from ~105 to 125 ka (thousand years ago), from Lingjing, Xuchang, China, which are referred to as “Late Pleistocene archaic human crania” by the authors.21 The cranial capacity of one cranium (Xuchang 1) is estimated at 1,800 cc, considerably above the modern human mean of 1,345 cc, and exhibits Homo erectus features, such as prominent brow ridge, a nuchal torus (although small), low vault height, and the widest point of the cranium being low.21 According to Roberto Sáez, the Xuchang skulls were initially considered Homo erectus.22 On the Xuchang crania Ann Gibbons writes:
“Nor are the new fossils late-occurring representatives of other archaic humans such as H. erectus or H. heidelbergensis, two species that were ancestral to Neandertals and modern humans. The skulls are too lightly built and their brains are too big, according to the paper.”23
In her brief article, Gibbons discusses the possibility that the crania belong to the mystery species, the Denisovans. But is ruling out Homo erectus a bit hasty? Another relevant question is whether the Denisovans were indeed Homo erectus, but that is not the focus here. Thickness of the cranial vault is arguably best measured by the thickness at the parietal eminence (a prominence near the centre of the external surface of the parietal bone), and this measurement in Xuchang 1 (7.9 mm24) is still within the lower range (7.5 mm) of Homo erectus measurements by Kennedy.25 Although this measurement was not given for the newly described Homo erectus crania, the vault thicknesses at another point on the cranium, called the bregma (junction point of the coronal and sagittal sutures), were listed as 7 mm and 8 mm for the DAN5/P1 and BSN12/P1 crania, respectively,26 compared to 8 mm at the bregma for the Xuchang 1 cranium.27 Being ‘lightly built’ would involve thin cranial vault bones, but the thickness of the vault bones appear to be still within the Homo erectus range for the Xuchang 1 cranium, as indicated above, along with the other erectus-like features. This indicates that it may essentially only be large brain size and changes to the form of the braincase (likely related to brain expansion) that rule out the Xuchang 1 cranium from being assigned to Homo erectus.
If crania are ruled out from belonging to Homo erectus essentially because of a large cranial capacity, then of course one will end up with a ‘species’ of only small-brained individuals. As such, and as a perfect example of faulty logic and circular reasoning, after arbitrarily ruling out crania with large cranial capacities as belonging to Homo erectus, some evolutionists then use this as proof that there are no Homo erectus crania with large cranial capacities.

Also, the boundary between Homo erectus and Homo heidelbergensis crania (figure 3) appears blurry, with cranial capacity an important factor influencing taxonomic status.28 Therefore, Homo erectus and Homo heidelbergensis should not be regarded as separate ‘species’. The cranial capacity range of 30 fossil specimens assigned to Homo heidelbergensis sensu lato is between 1,057.5 and 1,450 cc (the mean is about 1,244 cc).29
Considering the above factors, it does not appear correct to conclude that the cranial capacities of all Homo erectus crania are below the modern human mean, nor that a smaller cranial capacity correlates with greater robusticity. In this regard, it should be pointed out that the smaller Gona DAN5/P1 cranium was stated by Semaw et al. as being “more gracile”, whereas the larger Gona BSN12/P1 cranium was described as “more robust”.19 To explain the difference in size and morphology between the Gona specimens, the authors suggested that the older DAN5/P1 individual may have retained “more primitive anatomy (e.g. smaller size, gracile vault, and thin supraorbital tori) than the younger BSN12/P1 fossil”, or that the variation between the two specimens was “primarily a consequence of sexual dimorphism within a single species”, or that the “fossils might reflect a degree of taxonomic diversity previously unrecognized in the Afar for the genus Homo”.19
Small cranial capacity in Homo erectus

An important question is why so many of the Homo erectus crania have a small cranial capacity. Some specimens with small cranial capacities, such as those from Dmanisi, Georgia, in particular Skull 5 (figure 4), consisting of the cranium D4500 and the lower jaw D2600,30 can possibly be explained by pathology, such as cretinism, in some members of a local ‘Homo erectus’ population.13 With a cranial capacity of 546 cc, the adult D4500 cranium has the smallest cranial capacity of the Dmanisi sample (and any adult Homo erectus specimen known), although the cranial capacities of the other four Dmanisi crania (D2280: 730 cc; D2282: 650–660 cc; D2700: 600 cc; D3444: 625 cc)31 are also well below the average for Homo erectus. Cretinism (congenital hypothyroidism) “can reduce brain size by approximately 50%”,32 and so would be one possible explanation for the low cranial capacity observed in some of the Dmanisi crania. The very small brain size, as inferred by the small cranial capacities (brain size and cranial capacity are positively correlated), and the variability in craniomandibular morphology seen in specimens from the Dmanisi sample, including the odd-looking Skull 5, indicate something abnormal about this population. Similarly, the Homo naledi fossil sample from South Africa may, rather than being a new ‘species’, instead consist of ‘erectus-like’ humans, some of whom suffered from cretinism (explaining some of the strange morphology).33
However, while some small-brained Homo erectus specimens can possibly be explained as a result of pathology, there are too many specimens with small cranial capacity for pathology to be an explanation for all of them. And even if some may have been pathological, it is more likely that they were, for example, pathological Homo erectus-type humans, not pathological anatomically modern humans. The most logical conclusion appears to be that most Homo erectus specimens were not pathological and that there was a huge natural variation in cranial capacity of these humans, particularly if you include Homo heidelbergensis and other specimens (even the Neandertals) as part of this group (I prefer to call them ‘robust humans’). For reasons unknown many members of this group had small brain size, and hence small cranial capacity.
Small cranial capacity in anatomically modern humans
Is it possible that the newly described Gona DAN5/P1 cranium, with an estimated cranial capacity of ~598 cc, was of a Homo erectus individual with ‘normal’ human intelligence? Given that the DAN5/P1 cranium was directly associated with both simple Oldowan and more complex Acheulian tools, this is a very important question.
According to Kliemann et al., “Patients who had hemispherectomy in childhood may retain surprisingly high levels of cognitive and sensorimotor abilities.”34 Hemispherectomy is the surgical removal of one cerebral hemisphere (half of the brain). The authors also noted that “Language function has been studied in some detail, also showing near-complete recovery in many patients who had their language-dominant hemisphere resected [cut out].”35 Hence, this indicates that a large brain mass is not essential in order to have relatively ‘normal’ intelligence.
As the brain size or cranial capacity (and intelligence) of only a miniscule fraction of the billions of anatomically modern humans that exist (or have existed) have been measured or estimated, it is difficult to know what the lower limit of brain size for ‘normal’ intelligence is. One of the smallest brain sizes documented of a modern human with normal intelligence was from Daniel Lyon, a man of small stature (height of 1.55 m; weight 65.8 kg), with a brain that weighed 680 grams, yielding a brain volume of 624 cc, as calculated by John Skoyles.36 Using formulae linking brain volume or brain mass to cranial capacity, the cranial capacity of Lyon’s cranium can be estimated to be about 660 cc or 694 cc, depending on the method used.13 Perhaps the mean (677 cc) of the two values is the most reasonable estimate. This indicates, at least in theory, that most of the small-brained Homo erectus specimens, possibly including the individual associated with the DAN5/P1 cranium, could have been people with ‘normal’ intelligence.
The form of the Homo erectus cranium
A subject of importance to the creation model is explaining why the skeletons, particularly the skulls, of humans such as Homo erectus (as well as those classified as Neandertals and Homo heidelbergensis), were so rugged or robust compared to anatomically modern humans. Some creationists see environmental factors and genetic effects, such as the founder effect, genetic drift, and inbreeding, as most likely sufficient to explain the differences in skeletal features between modern humans and so-called cavemen.37 It has been proposed that there was a period of rapid human diversification beginning during the construction of Babel, resulting in extreme morphological variability in post-Babel humans repopulating the earth.38,39 Other creationists believe the stark difference in morphology between robust and anatomically modern humans warrants an explanation per se, that is, not just an acknowledgment that there was considerable variation in features, but why.
As such, it has been suggested that Homo erectus, Neandertals and the like all arose via ‘devolution’ from anatomically modern post-Babel populations—from the genetic consequences of inbreeding, drift, and mutational effects, after splitting off from anatomically modern humans.40 However, this would have to have happened independently, in a relatively short time, in many different parts of the world, which seems a little too coincidental to be realistic.
Along with a number of other creationists who have contemplated this issue, I consider the robust human forms, such as those with the features observed in the typical Homo erectus and Neandertal remains, to in general be non-pathological. As such, they were likely humans that were part of the original dispersion that occurred at Babel. Hence, an explanation that is worth considering is that the robust morphology observed in these humans is linked to longevity. According to creationist Carl Wieland:
“The extinction of human lines with more robust morphology (Neanderthal, erectus) may correlate with extinction of longevity. The robusticity may be the result of genetic longevity/delayed maturation or the same populations may have had [possibly linked] genes for longevity and robusticity.”41
Hence, one explanation why humans such as Homo erectus, Homo heidelbergensis, and Neandertals, had different morphology compared to modern humans, particularly in the skull, is that it could reflect changes in development of these early post-Flood individuals, compared to modern humans, possibly linked to longevity.42 A factor that cannot be ignored in a creationist model of human origins is the biblical long lifespans of individuals in the pre-Flood world, and to some extent the early post-Flood earth, as derived from Old Testament records. In the pre-Flood world lifespans of around 900 years appear to have been common, and even individuals born early post-Flood (within a few hundred years of the Flood having occurred) are recorded as having lived for hundreds of years.43,44
If initially (after the Fall) humans were designed to live for hundreds of years, then this would most likely have a bearing on development processes and timings. Simply put, longevity would probably be associated with changes in development, not just the aging process. Having thickened cranial vault bones, a heavily built face, thick-boned jaws, and thick postcranial bones may have been necessary, or at least an advantage, for the body to cope with these long lifespans.
If longevity was linked to development processes associated with the above-mentioned robusticity, then robust features would be expected to disappear with shorter lifespans, although, as Wieland notes, “some of the genes coding for some of their distinctive bony anatomy were apparently passed on to some of today’s populations”.45 Hence, robust features do not correlate with the potential for longevity in recent times. A genetic mechanism for robusticity would possibly involve the control of hormones related to bone growth and maintenance. According to Susan Crockford, an evolutionary expert in this area, the
“… distinctive skeletal morphology possessed by Neandertals is almost certainly the result of a pattern of thyroxine secretion (and, therefore, of prenatal and postnatal growth rates) that differed markedly and consistently from that of modern humans. These Neandertal traits may resemble superficially the pathological changes associated with congenital iodine deficiency because they reflect different amounts of thyroxine available for skeletal growth at particular stages of development as compared with healthy modern humans.” 46
Thyroxine (T4) is an important thyroid hormone, and a certain thyroid hormone secretion pattern could have existed in people with robust skeletal morphology, such as Homo erectus and the Neandertals, linked to the genetic mechanism for long lifespans. Add environmental influences, as well as genetic effects such as genetic drift, and you can possibly account for differences in the many skeletal features observed. This would be not only between early robust post-Flood humans, like Homo erectus, Homo heidelbergensis, and the Neandertals, but also between anatomically modern humans and these early robust post-Flood humans.
Explaining the skeletal form, particularly the crania, of robust humans, such as Homo erectus, is very much a work in progress. It is hoped that further work in this area will yield more answers or clues as to why these robust humans possessed crania that appear rugged and/or robust compared to the relatively gracile crania typical of anatomically modern humans.
Conclusions
The recent paper describing two new Homo erectus crania directly associated with both Oldowan and Acheulian stone tools raises issues regarding Homo erectus that are of great interest to the creation model of human origins. The latest evidence indicates that humans classified as Homo erectus made both Acheulian and Oldowan stone tools, although the simplicity of making the latter indicates that some Oldowan tools could possibly have been by-products of manufacturing the Acheulian tools. Also, the phantom species Homo habilis (most of whose members should be reassigned to the australopithecines) was likely not the maker of Oldowan tools. Although a high proportion of Homo erectus crania have small cranial capacities, the robust nature of Homo erectus crania is not necessarily linked to small brain size, as Homo erectus appears to be a species by and large arbitrarily defined as having a small cranial capacity. If other robust human crania are included in the Homo erectus group, like those belonging to Homo heidelbergensis, the Xuchang crania, and even the Neandertals, then the correlation between cranial robusticity and cranial capacity disappears. Why there is a high proportion of robust Homo erectus crania with small cranial capacities is unclear, but information from recent anatomically modern humans indicates that most of the small-brained Homo erectus individuals could, at least in theory, have possessed ‘normal’ intelligence. Why the robust cranial form seen in Homo erectus is different from that in anatomically modern humans is a matter of debate, with more work needed to be done in this area.
References and notes
- Semaw, S. et al., Co-occurrence of Acheulian and Oldowan artifacts with Homo erectus cranial fossils from Gona, Afar, Ethiopia, Science Advances 6(10):eaaw4694, pp. 1–8, 2020 | doi:10.1126/sciadv.aaw4694. Return to text.
- Semaw et al., ref. 1, supplementary file, pp. 17–18. Return to text.
- Semaw et al., ref. 1, pp. 1–2. Return to text.
- Semaw et al., ref. 1, p. 1. Return to text.
- Semaw et al., ref. 1, pp. 4–5. Return to text.
- Stringer, C. and Andrews, P., The Complete World of Human Evolution (revised edition), Thames & Hudson, London, p. 208, 2011. Return to text.
- Line, P., Homo habilis; in: Bergman, J., Line, P., Tompkins, J., Biddle, D, and Oard, M. (Eds.), The Apeman Controversy: A thorough examination of modern human evolution claims, Bartlett Publishing, Oklahoma City, OK, chap. 13, 2020 (in press). Return to text.
- Proffitt, T. et al., Wild monkeys flake stone tools, Nature 539:85, 2016. Return to text.
- Callaway, E., Monkey ‘tools’ raise questions over human archaeological record, 19 October 2016 | doi:10.1038/nature.2016.20816, nature.com/news/monkey-tools-raise-questions-over-human-archaeological-record-1.20816. Return to text.
- Schuster, R., Homo erectus was sexually dimorphic, new evidence shows, 4 March 2020, haaretz.com/archaeology/.premium-homo-erectus-was-sexually-dimorphic-new-evidence-shows-1.8627492. Return to text.
- Lewis, D., Skulls and skills varied in archaic Homo erectus, 6 March 2020, cosmosmagazine.com/palaeontology/skulls-and-skills-varied-in-archaic-homoerectus. Return to text.
- Stringer and Andrews, ref. 6, pp. 208–209. Return to text.
- Line, ref. 7, Homo erectus, chap. 14. Return to text.
- Vialet, A., Guipert, G., and Alçiçiek, M.C., Homo erectus found still further west: reconstruction of the Kocabaș cranium (Denizli, Turkey), C.R. Palevol 11:89–95, 2012. Return to text.
- Clarke, R.J., A corrected reconstruction and interpretation of the Homo erectus calvaria from Ceprano, Italy, J. Human Evolution 39:438–442, 2000. Return to text.
- Clarke, R.J., Australopithecus and Early Homo in Southern Africa; in: Delson, E. (Ed.), Ancestors: The Hard Evidence, Alan R. Liss, Inc., New York, p. 173, 1985. Return to text.
- Semaw et al., ref. 1, supplementary file, p. 3. Return to text.
- Semaw et al., ref. 1, p. 2. Return to text.
- Semaw et al., ref. 1, p. 3. Return to text.
- Molnar, S., Human Variation: Races, types, and ethnic groups, 6th edn, Pearson Prentice Hall, Upper Saddle River, New Jersey, p. 195, 2006. Return to text.
- Li, Z-Y. et al., Late Pleistocene archaic human crania from Xuchang, China, Science 355(6328):969, 2017. Return to text.
- Sáez, R., The Dali skull and other candidates to be Denisovan, 9 January 2019, nutcrackerman.com/2019/01/09/el-craneo-de-dali-y-otros-candidatos-a-denisovanos/. Return to text.
- Gibbons, A., Close relative of Neandertals unearthed in China, Science 355(6328):899, 2017. Return to text.
- Li et al., ref. 21, supplementary text, pp. 46 (figure S24), 62 (table S10). Return to text.
- Kennedy, G.E., On the autapomorphic traits of Homo erectus, J. Human Evolution 20:381–383, 1991. Return to text.
- Semaw et al., ref. 1, supplementary file, p. 40. Return to text.
- Li et al., ref. 21, supplementary text, p. 62 (table S10). Return to text.
- Line, ref. 7, Homo heidelbergensis, chap. 17. Return to text.
- Schoenemann, P., Hominid Brain Evolution; in: Begun, D.R. (Ed.), A Companion to Paleoanthropology, Wiley-Blackwell, West Sussex, UK, chap. 8, pp. 141–150, 2013; Arsuaga, J.L. et al., Neandertal roots: Cranial and chronological evidence from Sima de los Huesos, Science 344:1358–1363, 2014. See p. 59 of supplementary materials file for list of the cranial capacities of the Sima de los Huesos (SH) sample. 15 Homo heidelbergensis crania (excluding Sale as I have put that in the Homo erectus category) listed by Schoenemann and the 15 Homo heidelbergensis Sima de los Huesos (SH) crania measured/estimated by Arsuaga et al. were the 30 Homo heidelbergensis used in this analysis. Return to text.
- Lordkipanidze, D. et al., A Complete skull from Dmanisi, Georgia, and the evolutionary biology of early Homo, Science 342:326–331, 2013. Return to text.
- Rightmire, G.P. et al., Skull 5 from Dmanisi: Descriptive anatomy, comparative studies, and evolutionary significance, J. Human Evolution 104:52, 55, 2017. Return to text.
- Obendorf, P.J., Oxnard, C.E., and Kefford, B.J., Are the small human-like fossils found on Flores human endemic cretins? Proceedings of the Royal Society B 275:1287, 1290, 2008. Return to text.
- Line, P., Making sense of ‘Homo naledi’, Creation 40(4):36–38, 2018. Return to text.
- Kliemann, D. et al., Intrinsic functional connectivity of the brain in adults with a single cerebral hemisphere, Cell Reports 29:2398, 19 November 2019 | doi.org/10.1016/j.celrep.2019.10.067. Return to text.
- Kliemann et al., ref. 34, pp. 2398–2399. Return to text.
- Skoyles, J.R., Human Evolution Expanded Brains to Increase Expertise Capacity, not IQ, Psycoloquy 10(002), Brain Expertise (1), 1999. Return to text.
- Purdom, G., How are cavemen different? Answers 7(2):59, 2012. Return to text.
- Wood, T.C., An evaluation of Homo naledi and “Early” Homo from a young-age creationist perspective, J. Theology and Science Series B: Life Sciences 6:17–18, 23–25, 2016. Return to text.
- Wise, K.P., Paleontological note on Homo naledi, J. Creation Theology and Science Series B: Life Sciences 6:12–13, 2016. Return to text.
- Rupe, C. and Sanford, J., Contested Bones (2nd printing), FMS Publications, Longmeadow, MA, pp. 344–346, 2019. Return to text.
- Wieland, C., Decreased lifespans: have we been looking in the right place? J. Creation 8:141, 1994. Return to text.
- Line, P., Explaining robust humans, J. Creation 27(3):64–71, 2013. Return to text.
- Wieland, C., Living for 900 years, Creation 20(4):10, 1998. Return to text.
- Wieland, ref. 41, p. 139. Return to text.
- Wieland, ref. 41, p. 140. Return to text.
- Crockford, S.J., Commentary: Thyroid hormone in Neandertal evolution: A natural or pathological role? The Geographical Review 92(1):83, 2002. Return to text.



Readers’ comments
Comments are automatically closed 14 days after publication.