
Table des matières
Unité 1

Chapitre 1Argument : Le créationnisme est de la religion, pas de la science
Chapitre 2Argument : théorie de l’évolution et foi chrétienne sont compatibles
Chapitre 3Argument : La théorie de l’évolution est de la vraie science, et pas « juste une théorie »
Unité 2
Chapitre 4Argument : La sélection naturelle mène à la spéciation
Chapitre 5Argument : Certaines mutations sont bénéfiques
Chapitre 6Argument : La conception commune indique un ancêtre commun
Chapitre 7Argument : La « conception défectueuse » constitue un vestige de l’évolution
Chapitre 8Argument : Le registre fossile soutient la théorie de l’évolution
Unité 3
Chapitre 9Argument : Probabilité de l’évolution
Chapitre 10Argument : La « complexité irréductible »
Chapitre 11Argument : L’évolution du sexe
Chapitre 12Argument : L’évolution de l’humanité
Annexe 1Arguments courants en faveur de la théorie de l’évolution qui ont été rejetés
Annexe 2Arguments courants en faveur de la création à ne pas utiliser
« L’Évolution réfutée II » : chapitre 10
La suite de « L’Évolution réfutée » infirme les derniers arguments en faveur de la théorie de l’évolution (tels que présentés par PBS et Scientific American).
Argument : La « complexité irréductible »
Les évolutionnistes disent : « Les exemples de supposée “complexité irréductible” (comme l’œil, la si complexe cellule et le flagelle) peuvent trouver explication. »
Publié pour la première fois dans « Refuting Evolution 2 », chapitre 10.
Ce chapitre examine la réponse des évolutionnistes à l’argument de la « complexité irréductible » dans trois domaines : l’œil, la si complexe cellule et le flagelle. Scientific American présente le problème de la manière suivante :
14. Les êtres vivants ont des caractéristiques d’une complexité fantastique au niveau anatomique, cellulaire et moléculaire – qui ne pourraient pas fonctionner si elles étaient un tant soit peu moins complexes ou sophistiquées. La seule conclusion prudente est que ce sont les produits d’une conception intelligente, et non de l’évolution.
Cet « argument de la conception » est l’épine dorsale de la plupart des attaques récentes contre la théorie de l’évolution, mais c’est aussi l’un des plus anciens. En 1802, le théologien William Paley a écrit que si l’on trouve une montre de poche dans un champ, la conclusion la plus raisonnable est que quelqu’un l’a laissée tomber, et non que des forces naturelles l’ont créée à cet endroit. Par analogie, selon Paley, les structures complexes des êtres vivants doivent être l’œuvre d’une invention divine directe. Darwin a écrit « L’origine des espèces » pour répondre à Paley : il a expliqué comment les forces naturelles de sélection, agissant sur des caractéristiques héritées, pouvaient progressivement façonner l’évolution de structures organiques élaborées. [SA 83]
Effectivement, Gould, un expert de l’histoire de l’évolution, avait reconnu que Darwin écrivait pour contrer Paley. Une autre façon de dire qu’il avait des visées antithéistes,1 comme nous l’avons vu au chapitre 2. Cela n’empêche pas de nombreux universitaires chrétiens du dimanche de courber l’échine devant toutes les allégations de Darwin et de ses successeurs théophobes et qui, en retour, les considèrent avec autant de mépris que Lénine ses alliés « idiots utiles » à l’Ouest.2
L’œil aurait-il pu apparaître par évolution ?
Fait intéressant : l’œil, considéré par les évolutionnistes comme un exemple de « mauvaise conception » héritée de l’évolution (chapitre précédent), représente leur plus grand défi en tant qu’exemple de la superbe « complexité irréductible » de la création de Dieu. Scientific American dit :
Des générations de créationnistes ont tenté de contrer Darwin en citant l’exemple de l’œil comme une structure qui n’aurait pas pu apparaître par évolution. La capacité de l’œil à fournir la vision dépend de l’agencement parfait de ses parties, affirment ces critiques. La sélection naturelle ne pourrait donc jamais favoriser les formes transitionnelles nécessaires à l’évolution de l’œil – à quoi bon un demi-œil ? Anticipant cette critique, Darwin a suggéré que même des yeux « incomplets » auraient pu conférer des avantages (par exemple aider les créatures à s’orienter vers la lumière) et ainsi à survivre pour être davantage perfectionnées au cours de l’évolution. [SA 83]
Primo, cela ne tient pas compte de l’incroyable complexité ne serait-ce que du plus simple des organes photosensibles. Secundo, on ne saurait prétendre que la vision à 51 % aurait par nécessité un avantage sélectif suffisant par rapport à 50 % pour surmonter les effets de la dérive génétique, qui a tendance à éliminer même les mutations bénéfiques.3
La biologie a donné raison à Darwin : les chercheurs ont identifié des yeux primitifs et des organes photosensibles dans tout le règne animal et ont même retracé l’histoire de l’évolution des yeux grâce à la génétique comparée. (Il apparaît maintenant que dans diverses familles d’organismes, les yeux ont évolué indépendamment.) [SA 83]
Scientific American se contredit pour le coup. Si la génétique comparative a retracé l’histoire de l’évolution des yeux, comment se fait-il que ces derniers aient soi-disant évolué de manière indépendante ? En fait, les évolutionnistes reconnaissent que les yeux ont dû apparaître de façons indépendantes au moins 30 fois parce qu’il n’existe aucun modèle évolutif expliquant l’origine des yeux à partir d’un ancêtre commun. Cela signifie en réalité que, puisque les yeux ne peuvent avoir de parenté au moyen d’un ancêtre commun, qu’ils sont là et que seules les explications matérialistes comptent, passez muscade ! nous avons la preuve qu’ils ont évolué indépendamment les uns des autres !
Simulation de l’évolution de l’œil
PBS 1 se donne beaucoup de mal pour nous convaincre que l’œil a pu facilement évoluer. Dan Nilsson a expliqué une simulation informatique simpliste qu’il a publiée dans un article à grand tirage.4 S’inspirant de Darwin, qui a commencé par un endroit sensible à la lumière pour « expliquer » l’origine de l’œil, la simulation de Nilsson commence par une couche sensible à la lumière, avec un revêtement transparent à l’avant et une couche absorbant la lumière à l’arrière.
Voici comment se déroule la simulation. Primo, la couche sensible à la lumière se courbe peu à peu en forme de coupe, ce qui lui permet de percevoir de mieux en mieux la direction des rayons lumineux. Cela se poursuit jusqu’à ce qu’elle devienne un hémisphère rempli de substance transparente. Secundo, le rapprochement des extrémités, qui referme l’ouverture, augmente petit à petit la netteté de l’image, comme le fait un appareil photo à sténopé, parce qu’un trou plus petit coupe la lumière. Mais en raison de la diffraction de la lumière si le trou est trop petit, il y a une limite à ce processus. Ainsi, tertio, la forme et le gradient de l’indice de réfraction de la couverture transparente se transforment progressivement en une lentille à focalisation fine. Même si, par générosité, nous supposions que de telles simulations informatiques aient vraiment quelque chose à voir avec le monde réel de la biochimie, il y a de plus graves problèmes.
Cependant, le biochimiste Michael Behe a démontré que même un « simple » endroit photosensible a besoin d’un ensemble impressionnant de substances biochimiques au bon endroit et au bon moment pour fonctionner. Il affirme que chacune de ses « cellules fait paraître la complexité d’une moto ou d’un téléviseur dérisoire en comparaison » et décrit une petite partie de ce qui entre en jeu :5
Au début, lorsque la lumière frappe la rétine, un photon interagit avec une molécule appelée 11-cis-rétinal, qui se réarrange en quelques picosecondes en trans-rétinal. (Une picoseconde [
10-12 sec ] correspond à peu près à la durée nécessaire à la lumière pour parcourir la largeur d’un cheveu humain.) Le changement de forme du rétinal entraîne celui d’une protéine, la rhodopsine, en lien étroit avec celui-ci. La métamorphose de la protéine modifie son comportement. Désormais appelée métarhodopsine II, elle se colle à une autre protéine, la transducine. Avant de se heurter à la métarhodopsine II, la transducine a étroitement lié une petite molécule appelée GDP. Mais lorsque la transducine interagit avec la métarhodopsine II, le GDP se détache et une molécule appelée GTP se lie à la transducine (le GTP a une parenté étroite avec le GDP, mais il en est différent.)La GTP-transducine-métarhodopsine-II se lie maintenant à une protéine appelée phosphodiestérase, située dans la membrane interne de la cellule. Une fois fixée à la métarhodopsine II et à son entourage, la phosphodiestérase acquiert la capacité chimique de « couper » une molécule appelée GMPc (un parent chimique du GDP et du GTP). Au départ, il y a beaucoup de molécules de GMPc dans la cellule, mais la phosphodiestérase en réduit la concentration, tout comme une bonde tirée fait baisser le niveau de l’eau dans une baignoire.
Il y a de même bien plus de difficulté à obtenir une couche transparente que ne le pensent les chercheurs. La meilleure explication de la transparence de la cornée repose sur la théorie de la diffraction, qui démontre que la lumière ne se disperse pas si l’indice de réfraction reste constant sur des distances supérieures à la moitié de la longueur d’onde de la lumière. Cela exige une organisation très précise des fibres de la cornée, ce qui nécessite des pompes chimiques complexes pour garantir l’exactitude de la teneur en eau.6
Par conséquent, ces simulations ne partent pas d’un état simple mais présupposent une grande complexité dès le départ. Dans leur article initial, les chercheurs admettent pareillement qu’« un œil n’a guère de sens en soi », car la capacité de percevoir la lumière ne sert à rien si l’organisme manque d’une machinerie informatique sophistiquée pour utiliser cette information. Par exemple, il doit avoir la capacité de traduire « atténuation de l’intensité des photons » en « l’ombre d’un prédateur en est responsable » en « je dois prendre des mesures d’évitement », et pouvoir agir selon ladite information pour qu’elle présente une quelconque valeur sélective. De même, la courbure du début, avec sa modeste capacité à détecter la direction de la lumière, ne fonctionnerait que si la créature disposait du « logiciel » approprié pour l’interpréter. La perception d’images réelles présente encore plus de difficultés. Les personnes qui retrouvent la vue après des années de cécité ont besoin d’un certain temps pour apprendre à voir correctement. Il convient de noter qu’une grande partie du traitement de l’information a lieu dans la rétine avant que le signal n’atteigne le cerveau.
Le fait de montrer une série d’yeux plus complexes dans la nature et d’affirmer qu’il s’agit d’une séquence évolutive constitue de même un sophisme. C’est comme si l’on classait un certain nombre de types d’avions par ordre de complexité et que l’on affirmait ensuite que les avions simples ont évolué vers des avions complexes, au lieu d’avoir été conçus. D’une part, les yeux ne peuvent descendre d’autres yeux à proprement parler : ce sont les organismes qui transmettent les gènes des yeux à leurs descendants. Un point important si l’on considère l’œil du nautile, un appareil photo à sténopé. Il ne saurait s’agir d’un ancêtre de l’œil des vertébrés, car le nautile, d’une manière générale, n’est pas un ancêtre des vertébrés, même d’après les évolutionnistes !
Les moteurs rotatifs du flagelle bactérien
Scientific American cite un autre exemple difficile de complexité irréductible, les moteurs rotatifs du flagelle bactérien, mais n’apporte aucune véritable réponse.
15. Des découvertes récentes prouvent que même au niveau microscopique, la vie possède une dimension de complexité qui n’a pas pu naître de l’évolution.
La « complexité irréductible » est le cri de guerre de Michael J. Behe de l’université de Lehigh, auteur de « Darwin’s Black Box: The Biochemical Challenge to Evolution ». Pour illustrer la complexité irréductible, Behe choisit un exemple du quotidien : la souricière, une machine qui ne pourrait pas fonctionner si l’une de ses pièces manquait et dont les pièces n’ont de valeur qu’en tant que parties de l’ensemble.
Ce qui est vrai pour la souricière l’est encore plus pour le flagelle bactérien, un organite cellulaire en forme de fouet utilisé pour la propulsion et qui fonctionne comme un moteur hors-bord. Les protéines qui composent le flagelle sont étonnamment bien agencées pour former des composants de moteur, un joint universel et d’autres structures comme celles qu’un ingénieur humain pourrait spécifier. Selon Behe, la possibilité que cet ensemble complexe soit né de modifications évolutives est pratiquement nulle, ce qui est le signe d’une conception intelligente. [SA 84]
Absolument (voir diagramme ci-dessous).
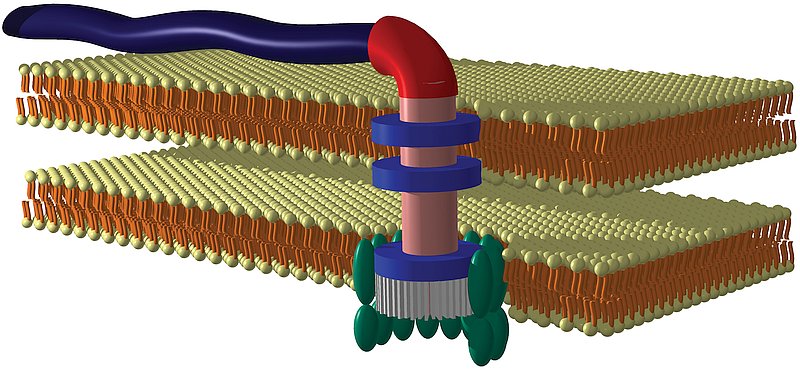
Flagelle bactérien avec moteur rotatif, présentant les caractéristiques suivantes :
- Auto-assemblage et réparation
- Moteur rotatif à refroidissement par eau
- Système d’entraînement par force motrice protonique
- Marche avant et marche arrière
- Vitesses de fonctionnement jusqu’à 100 000 tr/min
- Capacité d’inversion de sens au quart de tour
- Système de transduction des signaux câblé avec mémoire à court terme
[extrait de « Bacterial Flagella: Paradigm for Design », vidéo,
www.arn.org/news-videos/videos.html]
Il soulève des points similaires concernant le mécanisme de coagulation du sang et d’autres systèmes moléculaires.
Pourtant, les biologistes évolutionnistes ont des réponses à ces objections. Tout d’abord, il existe des flagelles de formes plus simples que celle citée par Behe, il n’est donc pas nécessaire que tous ces composants soient présents pour qu’un flagelle fonctionne. Les composants sophistiqués de ce flagelle ont tous des précédents ailleurs dans la nature, comme l’ont décrit Kenneth R. Miller de l’université Brown et d’autres. [SA 84]
Miller ne représente guère l’incarnation de la fiabilité. Behe a en outre répondu à des critiques comme celui-ci.7
En fait, l’ensemble de l’assemblage du flagelle est extrêmement similaire à un organite que Yersinia pestis, la bactérie de la peste bubonique, utilise pour injecter des toxines dans les cellules. [SA 84]
Cela provient en réalité de l’utilisation abusive par le National Center for Science Education des recherches du Pr Scott Minnich, généticien et professeur agrégé de microbiologie à l’université de l’Idaho. Cet expert de renommée mondiale sur le flagelle affirme que la croyance en la conception lui a apporté de nombreuses idées de recherche. Ses travaux de recherche démontrent que le flagelle ne se forme pas à plus de 37° C et qu’à la place, certains organites sécrétoires se forment à partir du même ensemble de gènes. Mais cet appareil sécrétoire, ainsi que l’appareil de forage de la bactérie de la peste, correspondent à une dégénérescence du flagelle, qui, selon Minnich, a fait apparition le premier, bien qu’il soit plus complexe.8
Le point clé est que les structures constitutives du flagelle, dont Behe suggère qu’elles n’ont aucune valeur en dehors de leur rôle dans la propulsion, peuvent remplir de multiples fonctions qui auraient contribué à favoriser leur évolution. [SA 84]
En fait, ce que Behe dit vouloir dire par complexité irréductible, c’est que le flagelle ne pourrait pas fonctionner sans une quarantaine de composants protéiques tous organisés de la bonne manière. L’argument de Scientific American revient à prétendre que si les composants d’un moteur électrique existent déjà dans un garage à voitures électriques, ils pourraient s’assembler d’eux-mêmes pour former un moteur fonctionnel. Cependant, la bonne organisation revêt la même importance que les bons composants.
L’évolution finale du flagelle pourrait donc n’avoir impliqué que la nouvelle recombinaison de parties sophistiquées qui ont initialement évolué à d’autres fins. [SA 84]
Minnich souligne que seuls dix des quarante composants peuvent s’expliquer par la cooptation, mais que les trente autres sont entièrement nouveaux. En outre, le processus même d’assemblage dans la bonne séquence nécessitant d’autres mécanismes de régulation, il s’avère donc irréductiblement complexe en soi.9
La coagulation sanguine
Scientific American cite un autre problème sérieux pour la théorie de l’évolution : la coagulation sanguine.
De même, le système de coagulation du sang semble impliquer la modification et l’élaboration de protéines qui servaient à l’origine à la digestion, selon les études de Russell F. Doolittle de l’université de Californie à San Diego. Ainsi, une partie de la complexité que Behe qualifie de preuve de dessein intelligent n’est pas du tout irréductible. [SA 84]
Il s’agit une fois de plus de hâblerie de la part de l’athée Doolittle, ou tout du moins de mauvaise compréhension de texte. Il a cité des expériences récentes montrant que des souris pouvaient survivre malgré l’élimination de deux des composants de la cascade de coagulation du sang (le plasminogène et le fibrinogène). Cela aurait soi-disant démontré que la cascade actuelle n’est pas irréductiblement complexe mais clairement réductiblement complexe. Mais l’expérience a en réalité montré que les souris dépourvues des deux composants se portaient mieux que celles privées uniquement de plasminogène, car ces dernières souffraient de caillots non éliminés. Mais les premières ne sont pas aussi saines que Doolittle l’a laissé entendre, car la seule raison pour laquelle elles ne souffrent pas de caillots non nettoyés est qu’elles n’ont pas de système de coagulation fonctionnel du tout ! Un système de coagulation non fonctionnel (bien qu’il possède les nombreux composants restants) ne constitue pas un intermédiaire évolutif que la sélection naturelle pourrait affiner pour produire un système de coagulation adéquat. Cette expérience prouve plutôt l’inverse, car la sélection naturelle éliminerait l’étape suivante (c’est-à-dire le passage de l’absence de plasminogène et de fibrinogène au seul fibrinogène) en raison des caillots non nettoyés.10
Une complexité d’un autre type – la « complexité spécifique » – est la pierre angulaire des arguments en faveur de la conception intelligente avancés par William A. Dembski, de l’université Baylor, dans ses livres « The Design Inference » et « No Free Lunch ». En substance, son argument est que les êtres vivants sont complexes d’une manière que des processus non dirigés et aléatoires ne pourraient jamais produire. La seule conclusion logique, affirme Dembski, en écho à Paley il y a deux siècles, est qu’une intelligence surhumaine a créé et façonné la vie.
L’argument de Dembski comporte plusieurs lacunes. Il est faux d’insinuer que le champ des explications ne comprend que des processus aléatoires ou des intelligences conceptrices. Les chercheurs en systèmes non linéaires et en automates cellulaires de l’Institut de Santa Fe et d’ailleurs ont démontré que des processus simples et non dirigés peuvent produire des motifs extraordinairement complexes. Une partie de la complexité observée dans les organismes peut donc émerger de phénomènes naturels que nous comprenons encore à peine. Mais cela est très différent de l’affirmation selon laquelle la complexité n’a pas pu apparaître naturellement. [SA 84]
Ça, c’est de la foi aveugle ! Mais dans la pratique, comme le souligne Dembski, la complexité spécifiée sert, dans tous les cas de figure sauf en biologie, de preuve en faveur de la conception, y compris dans la recherche d’une intelligence extraterrestre [allusion au SETI (Search for Extraterrestrial Intelligence – Recherche d’Intelligence Extraterrestre – NdT), programme de la NASA qui consiste à scanner les fréquences venues de l’espace pour y trouver des motifs provenant de civilisations extraterrestres]. La complexité biologique constituant la seule exception proposée par les évolutionnistes, cela empeste le plaidoyer pro domo.11
Outre l’œil humain, le flagelle et la coagulation sanguine, il existe une multitude d’autres exemples de complexité irréductible dans la nature. J’ai déjà fait allusion au mécanisme dynamique de collage des pattes des insectes. Les pattes collantes des geckos illustrent elles aussi à merveille l’ingéniosité de Dieu.12 Les découvreurs évolutionnistes de sa structure la décrivent comme « dépassant les limites de la technologie humaine. »13 Parmi les autres exemples de conception, citons les yeux du homard et leur géométrie réfléchissante carrée unique, qui a inspiré des télescopes à rayons X avancés et des producteurs de faisceaux,14 ainsi que le moteur de l’ATP synthase..
Références et notes
- Wieland, C., « Darwin’s real message: have you missed it? » Creation 14(4):16–19, septembre 1992 ; J. Sarfati, analyse de K. Birkett, « The Essence of Darwinism » ; cf. « Evangelical compromise misses the essentials ». Revenir à l’article.
- Sarfati, J., « The Skeptics and their ‘Churchian’ allies ». Revenir à l’article.
- Voir mon traitement du sujet de l’évolution de l’œil dans « Stumbling over the impossible: refutation of Climbing Mt Improbable », Journal of Creation 12(1):29–34, 1998 ; cf. « Eye evolution, a case study ». Revenir à l’article.
- Nilsson, D. E. et Pelger, S., « A pessimistic estimate of the time required for an eye to evolve », Processings of the Royal Society of London B 256:53–58, 1994. Revenir à l’article.
- Behe, M. J., « Darwin’s Black Box: The Biochemical Challenge to Evolution », The Free Press, New-York, district de New-York, 1996, p. 46. Revenir à l’article.
- Gurney, P. W. V., « Dawkins’ eye revisited », Journal of Creation 15(3):92–99, 2001. Revenir à l’article.
- « Behe responds to various critics », www.trueorigin.org/behe08.asp. Revenir à l’article.
- Cf. Scott Minnich, « Bacterial flagella: spinning tails of complexity and co-option », www.idurc.org/yale-minnich.html, disponible sur web.archive.org. Revenir à l’article.
- Unlocking the Mystery of Life, vidéo, Illustra Media, 2002. Revenir à l’article.
- Pour plus d’informations, cf. « In defense of the irreducibility of the blood clotting cascade », par Behe, www.trueorigin.org/behe03.asp. Revenir à l’article.
- Grigg, R., « A brief history of design », Creation 22(2):50–53, mars 2000. Revenir à l’article.
- Sarfati, J., « Great gecko glue? », Creation 23(1):54–55, décembre 2000. Revenir à l’article.
- Autumn, K., et al., « Adhesive force of a single gecko foot hair », Nature 405(6787): 681–685, 8 juin 2000 ; perspective par H. Gee, « Gripping feat », même numéro, p. 631. Revenir à l’article.
- Sarfati, J., « Lobster eyes—brilliant geometric design », Creation 23(3)12–13, juin 2001. Revenir à l’article.
Note sur les citations : Les citations de l’article de John Rennie dans Scientific American seront marquées « SA », suivi du numéro de page. Les citations et autres mentions de la série télévisée « Evolution » de PBS-TV porteront la mention « PBS », suivie du numéro de l’épisode ; par exemple, « PBS 6 » fait référence à l’épisode 6. Revenir à l’article.
Readers’ comments
Comments are automatically closed 14 days after publication.